| |

Bacteria CellWhat is a Bacteria ?The prokaryotic group of unicellular creatures, which includes bacteria, lacks a few organelles and a real nucleus. The Cellular Structure of a BacteriumBacteria are noted for having straightforward body plans. Bacteria are prokaryotic creatures because they are single-celled microorganisms without a nucleus or other cell organelles. 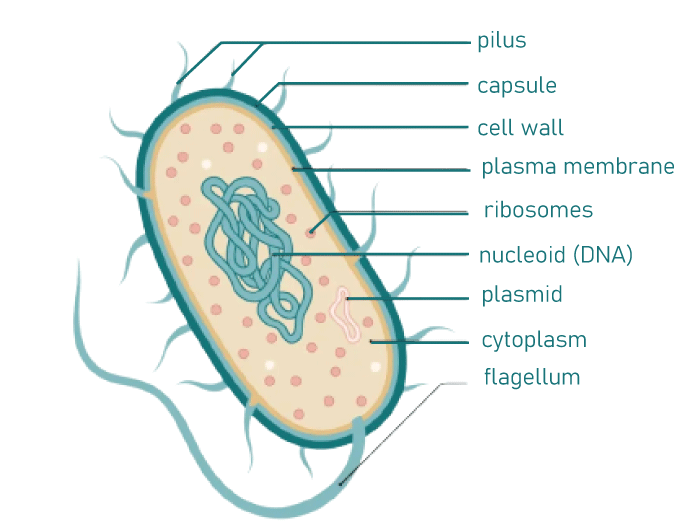
They are also exceptionally adaptable organisms, able to endure hostile environments, and also known as Extremophiles organisms. Extremophiles are further divided into several categories according to the habitats they live in:
The protective cell wall of bacteria, composed of a unique protein called peptidoglycan, is another intriguing aspect of bacteria. The elements of the bacterial cell wall provide a crucial foundation for dividing bacteria. The only other place this specific protein may be found is in the bacterial cell walls. Yet, only a small percentage of them lack this cell wall, and others have an additional layer of defense known as a capsule. One or more flagella or pili are attached to the outer layer and serve as a locomotory organ. Certain bacteria may be assisted in adhering to the cells of the host by pili. Except for ribosomes, they lack any organelle found in an animal or plant cell. Protein synthesis takes place in ribosomes. They also possess a second circular piece of DNA known as a plasmid. Certain bacterial strains become resistant to antibiotics thanks to these plasmids. Reproduction of BacteriaBinary fission is the asexual reproductive strategy used by bacteria. A bacterium splits into two daughter cells after dividing. Both to each other and to the parent cell, these are identical. The fission process begins with DNA replication within the parent bacterium. Cell eventually grows longer and divides into two daughter cells. Reproduction occurs at different rates and times depending on environmental factors including temperature and nutrition availability. When the conditions are right, Escherichia coli or E. coli produces roughly 2 million bacteria every seven hours. Although bacterial reproduction is purely asexual, it can occasionally engage in sexual reproduction. Bacterial genetic recombination may take place via conjugation, transformation, or transduction. Due to genetic variation in these situations, the bacteria may develop antibiotic resistance (as opposed to asexual reproduction, where the same genetic material is present in generations). Useful BacteriaNot all microorganisms pose a threat to people. Several microorganisms are advantageous in various ways. Here are a few advantages of bacteria:
Harmful BacteriaSome microorganisms have a wide range of disease-causing potentials. They are to blame for a number of infectious diseases, including tooth decay, tuberculosis, diphtheria, syphilis, and pneumonia. Antibiotics and other recommended medications can be used to treat their effects. Yet, precaution is considerably more effective. By sterilizing or disinfecting exposed surfaces, instruments, tools, and other utilities, the majority of these disease-causing germs can be wiped off. These techniques include heat, cleaning agents, UV rays, pasteurization, boiling, and others. Cell MorphologyBacterial morphology may be the most fundamental structural characteristic (shape). Some of its typical illustrations include:
A given bacterial species' cells typically have a certain shape. However, growing conditions can change this. Certain bacteria, such as Caulobacter, have elaborate structures that contain reproductive spores and intricate life cycles that involve the creation of stalks and appendages (e.g., Myxococcus, Streptomyces). When grown on Petri plates, a bacteria typically produce different colony morphologies and different cell morphologies when observed under a light microscope. With some exceptions, the size of the bacteria is their most glaring structural feature. For instance, the "average" sized bacteria Escherichia coli cells are around 2 m (micrometers) long, 0.5 m in diameter, and have a cell volume of 0.6-0.7. It produces roughly 1 g of wet cell mass (109 cells/ml). Compact size is crucial because it enables a high surface area-to-volume ratio that quickly promotes nutrition intake, intracellular distribution, and waste excretion. The diffusion of nutrients and waste products over the bacterial cell membrane reduces the rate at which the microbial metabolism may occur at low surface area-to-volume ratios, rendering the cell less evolutionary suitable. Although the exact cause of giant cells is unknown, the extra volume is assumed to be mostly employed to store some more nutrients. Cell WallThe cell wall and membrane together make up the cell envelope. The bacterial cell wall gives the cell structural integrity, just like in other species. The main purpose of the cell wall in prokaryotes is to shield the cell from internal turgor pressure, which is brought on by the considerably higher concentrations of proteins and other molecules inside the cell than in the surrounding environment. The peptidoglycan, found just outside of the cell membrane, sets the bacterial cell wall apart from that of all other organisms. The polysaccharide backbone of peptidoglycan is composed of equal numbers of N-acetylglucosamine (NAG) and N-acetylmuramic acid (NAM) residues alternated. Bacterial cell walls are stiff, and the shape of the cells is always determined by peptidoglycan. Because of its porous nature, it is not thought to be a permeability barrier for tiny substrates. Peptidoglycan is a component of all bacterial cell walls, but not all have the same general structure (a few exceptions include extracellular parasites like Mycoplasma). As some eukaryotes lack the cell wall necessary for the bacterial survival, several antibiotics, most notably penicillins, and cephalosporins, terminate bacterial infections by disrupting the manufacture of cell walls without impacting human cells, which merely have a cell membrane. Gram-positive and gram-negative bacteria have various types of cell walls, which can be identified by their differences in how they react to the Gram stain. Particles that are as small as 2 nm can pass through the peptidoglycan for both bacteria types. A protoplast refers to a bacterial cell wall that has been completely destroyed, whereas a spheroplast refers to a bacterial cell wall that has only been partially removed. Penicillin and other beta-lactam antibiotics prevent the peptidoglycan cross-links from forming in bacterial cell walls. Human tears contain the lysozyme enzyme, which breaks down bacterial cell walls and serves as the body's primary defense against eye infections. Gram-positive cell wallThe peptidoglycan layer, also known as murein, makes up over 95% of the cell wall in some gram-positive bacteria. In comparison, it makes up only 5-10% of the wall in gram-negative bacteria. Gram-positive bacteria have thick cell walls. The gram-positive bacteria absorb the crystal violet dye, resulting in purple staining. Lysozymes can fully break down the cell walls of some gram-positive bacteria by attacking the links between N-acetylglucosamine and N-acetylmuramic acid. The walls of other gram-positive bacteria, such as Staphylococcus aureus, are resistant to lysozyme activity. On the carbon-6 of certain muramic acid residues, they have O-acetyl groups. Teichoic acids or polysaccharides may be the matrix components in the walls of gram-positive bacteria. The latter are relatively ubiquitous but have been detected only in gram-positive bacteria. Teichoic acids come in two main varieties: glycerol and ribitol teichoic acids. The latter is the more common. These acids are only found on the surface of many gram-positive bacteria and are polymers of glycerol phosphate and ribitol phosphate, respectively. Teichoic acid's precise role, however, is disputed and poorly understood. Lipoteichoic acid is a crucial component of the gram-positive cell wall. Its ability to serve as an antigenic is one of its goals. The membrane contains the lipid element, which helps the membrane hold it in place with the help of its adhesive capabilities. Gram-negative cell wallGram-negative cells have significantly thinner cell walls than gram-positive cells do. They also have a second plasma membrane located next to the cytoplasmic membrane and is superficial to their thin peptidoglycan layer. Gram-negative bacteria leave a pink stain on surfaces. Many of the antigenic traits of these strains are caused by the lipopolysaccharide's chemical makeup, which is frequently particular to certain bacterial subspecies. Plasma MembraneThe phospholipid bilayer that makes up the plasma membrane, also known as the bacterial cytoplasmic membrane, performs all the general functions of a cell membrane, including acting as a permeability barrier for the majority of molecules and acting as the site for the transport of molecules into the cell. Prokaryotic membranes serve as the site where a proton motive force is produced in addition to these roles in energy conservation. With a few exceptions, such as Mycoplasma and methanotrophs, bacterial membranes generally lack sterols in contrast to eukaryotes. Hopanoids, structurally similar substances found in many microorganisms, probably have the same purpose. Unlike eukaryotes, bacteria can have a wide array of fatty acids within their membranes. Hopanoids, structurally similar substances found in many microorganisms, probably have the same purpose. In contrast to eukaryotes, bacteria's membranes may contain a wide range of fatty acids. Bacteria can include fatty acids with extra methyl, hydroxy, or cyclic groups in addition to the usual saturated and unsaturated fatty acids. To maintain the ideal fluidity of the membrane, the bacterium can modify the relative proportions of these fatty acids (e.g., following temperature change). Mycobacteria and gram-negative bacteria both have an inner and an outer bacterial membrane. The lipid part of the bacterial outer membrane is impermeable to charged molecules because it is a phospholipid bilayer. Nonetheless, the outer membrane contains porin channels that enable the passive translocation of many ions, carbohydrates, and amino acids. As a result, these molecules can be found in the periplasm, which is the space between the cytoplasmic and outside membranes. The peptidoglycan layer and several proteins involved in substrate binding, hydrolysis, and extracellular signal receiving are found in the periplasm. Because of the substantial amounts of proteins and peptidoglycan present, the periplasm is assumed to exist in a gel-like condition as opposed to a liquid state. Signals received and substrates bound can be transmitted across the cytoplasmic membrane via transport and signaling proteins embedded there because of their placement between the cytoplasmic and outer membranes. Extracellular StructuresFimbriae and Pili Several Pseudomonadota members have protein tubes that protrude from the outer membrane known as fimbriae ("attachment pili"). They are frequently brief and abundantly distributed across the entire bacterial cell surface. Fimbriae typically make it easier for bacteria to connect to a surface or other cells to build a biofilm (e.g., animal cells during pathogenesis). Fimbriae are used by a few species (such as Myxococcus) for movement to aid in the development of multicellular structures like fruiting bodies. Pili are longer and sparsely distributed on the bacterial cell, despite having a structure that is comparable to fimbriae. Pili are known as "sex pili" or "conjugation pili" because they are involved in bacterial conjugation. Bacteria use Type IV pili (non-sex pili) to help them grip objects. Glycocalyx Glycocalyx is an extracellular polymer many bacteria release outside their cell walls. These polymers occasionally contain protein and typically contain carbohydrates. The largely impermeable architecture of capsules prevents them from being discolored by colors like Indian ink. These are structures that aid in preventing phagocytosis and desiccation from harming bacteria. The slime layer facilitates the formation of biofilms by bacteria on other cells or inanimate surfaces. Layers of slime may act as a cell's food store as well. Flagella Flagella are arguably the most identifiable extracellular bacterial cell structures. Flagella mediate bacterial motility, whip-like projections projecting from the bacterial cell wall. The bacterial species observed has a particular arrangement of flagella around its cell. Typical formats include:
Three fundamental parts comprise the bacterial flagellum: a whip-like filament, a motor complex, and a hook connecting them. The protofilaments, which each have thousands of flagellin subunits, make up the filament with a diameter of about 20 nm. The bundle may or may not be enclosed and is kept together by a cap. A proton-driven motor that propels rotation in the filament is located after a set of rings that anchor the flagellum in the inner and outer membranes. S-layers A cell surface protein layer called an S-layer, which also functions as the cell wall in some archaea, is known as the surface layer. All S-layers are crystalline and composed of a two-dimensional array of proteins, albeit the symmetry varies between species. Although their precise use is unknown, S-layers have been proposed to serve as a partial permeability barrier for large substrates. For instance, an S-layer might maintain extracellular proteins close to the cell membrane by obstructing their exocytosis. By protecting host defense mechanisms, an S-layer may aid in the facilitation of survival within the host in some pathogenic species. Intracellular StructureBacterial DNA and Plasmids Contrary to eukaryotes, bacteria have cytoplasm as their nucleus instead of a membrane-bound nucleus that houses their DNA. This indicates that the processes of translation, transcription, and DNA replication, which transfer cellular information, all take place in the same compartment and may interact with other cytoplasmic components, most notably ribosomes. Two sites can be found where bacteria have DNA:
In contrast to eukaryotes, which package their chromatin using histones, bacteria package their DNA into a highly compact supercoiled structure, the nature of which is still unknown. Despite a few cases of linear DNA, the majority of bacterial chromosomes are circular (e.g., Borrelia burgdorferi). Although some types of bacteria have been identified with several chromosomes, most bacteria only have one. In addition to chromosomal DNA, most bacteria also include tiny, autonomous DNA fragments called plasmids that frequently encode beneficial features but are not necessary for the survival of their bacterial host. Plasmids are simple for bacteria to acquire or lose, and they can be shared between bacteria as a means of horizontal gene transfer. Hence, plasmids in a bacterial cell can be thought of as extrachromosomal DNA. Ribosomes The ribosome, where all living things synthesize their proteins, is the internal structure found in the largest number in most bacteria. All prokaryotes have the 70S (S=Svedberg units) ribosomes, but the cytoplasm of eukaryotes has bigger 80S ribosomes. The 50S and 30S subunits make form the 70S ribosome. Whereas the 30S subunit contains the 16S rRNA, the 50S subunit contains the 23S and 5S rRNA. In eukaryotes, these rRNA molecules have different sizes and are complexed with several ribosomal proteins, which might differ slightly in number and type between different organisms. While the ribosome is the internal multiprotein complex in bacteria that is most frequently recognized, other big complexes do exist and can occasionally be observed using a microscope. Intracellular Membranes Certain germs have intracellular membranes in addition to (or as extensions of) their cytoplasmic membranes, which is uncommon for all bacteria. It was formerly thought that bacteria might have mesosomes-membrane folds-but further research revealed that they were simply byproducts of the chemicals used to prepare cells for electron microscopy. Nitrifying bacteria, methane-oxidizing bacteria, and phototrophs are a few types of bacteria that include internal membranes. Although these membranes more closely resemble organellar membranes in eukaryotes and are now of unclear function, intracellular membranes are also present in bacteria belonging to the poorly understood Planctomycetota group. Bacteria that use photosynthesis have intracellular membranes called chromophores. They include carotenoids and bacteriochlorophyll pigments, which are largely used for photosynthesis. Cytoskeleton The structural filaments in prokaryotes are collectively referred to as the prokaryotic cytoskeleton. Prokaryotic cells were once believed to lack cytoskeletons. However, improvements in imaging techniques and structural analysis have revealed the existence of filaments in these cells. Prokaryotes contain homologs for every significant cytoskeletal protein discovered in eukaryotes. In different prokaryotes, cytoskeletal components are crucial for cell division, protection, shape determination, and polarity determination. Nutrient storage structures The majority of bacteria do not always thrive in situations with abundant nutrition. Bacteria have a variety of ways to store nutrients for usage in times of scarcity in order to accommodate these sporadic levels of nutrients. For instance, many bacteria store extra carbon in the form of glycogen or polyhydroxyalkanoates. Some microorganisms store soluble nutrients such as nitrate in vacuoles. The most common form of sulfur storage is elemental granules, which can be deposited intra- or extracellularly. Sulfur granules are particularly prevalent in bacteria that use hydrogen sulfide as an electron source. Most instances mentioned below may be examined using a microscope and surrounded by a thin non-unit membrane to isolate them from the cytoplasm: 1. Inclusions: Included elements of the cell are those that do not have metabolic activity and are not surrounded by membranes. Crystals, pigments, lipid droplets, and glycogen are typical inclusions. Volutin granules are cytoplasmic inclusions of complex inorganic polyphosphate. Because of the metachromatic effect, these granules exhibit when stained with the blue dyes methylene blue or toluidine blue; they are known as metachromatic granules. 2. Gas vacuoles: Gas vacuoles are membrane-bound, spindle-shaped vesicles found in some planktonic bacteria and Cyanobacteria that provide buoyancy to these cells by decreasing their overall cell density. In order for the cells to maintain their position in the upper levels of the water column and continue to engage in photosynthesis, positive buoyancy is required. They consist of a protein shell with a very hydrophobic inner surface permeable to most gases but impervious to water (preventing water vapor from condensing inside). The hollow cylindrical shape of the gas vesicle makes it susceptible to collapsing when the atmospheric pressure rises. Natural selection has precisely honed the gas vesicle's structure to maximize its buckling resistance, including the externally reinforcing protein GvpC, which resembles the green thread in a braided hosepipe. There is a simple relationship between the diameter of the gas vesicle and the pressure at which it will collapse - the wider the gas vesicle, the weaker it becomes. Wider gas vesicles, as opposed to narrow gas vesicles, are more effective and offer greater buoyancy per unit of protein. Different species produce gas vesicles of varying diameters, enabling them to colonize the water column at various depths (fast-growing, highly competitive species with wide gas vesicles in the topmost layers; slow-growing, dark-adapted species with strong narrow gas vesicles in the deeper layers). Which species endure in various bodies of water depends on the diameter of the gas vesicle. The hydrostatic pressure produced by the complete water column is exposed to the cells in deep lakes that experience winter mixing. As a result, species with stronger, narrower gas vesicles will be selected. 3. Carboxysomes: Carboxysomes are bacterial microcompartments found in many autotrophic bacteria such as Cyanobacteria, Knallgasbacteria, Nitroso- and Nitrobacteria. The enzymes of carbon dioxide fixation in these species are found in these proteinaceous structures, which resemble phage heads in their appearance (particularly ribulose bisphosphate carboxylase/oxygenase, RuBisCO, and carbonic anhydrase). It is believed that carbon dioxide fixation can occur more quickly and effectively outside of the cytoplasm due to the high local concentration of enzymes and the quick conversion of bicarbonate to carbon dioxide by carbonic anhydrase. Certain Enterobacteriaceae are known to feature similar structures that house the coenzyme B12-containing glycerol dehydratase, a crucial enzyme in the conversion of glycerol to 1,3-propanediol (e. g., Salmonella). 4. Microcompartments: These proteinaceous structures, particularly ribulose bisphosphate carboxylase/oxygenase, RuBisCO, and carbonic anhydrase, are present in these species and resemble phage heads. The high local concentration of enzymes and the fast conversion of bicarbonate to carbon dioxide by carbonic anhydrase allow for more efficient and quick carbon dioxide fixing outside of the cytoplasm. The coenzyme B12-containing glycerol dehydratase, a vital enzyme in the conversion of glycerol to 1,3-propanediol, is known to be housed in structures that are identical in several Enterobacteriaceae (e.g., Salmonella). 5. Magnetosomes: Bacterial microcompartments called magnetosomes are present in magnetotactic bacteria, which enable them to sense and position themselves along a magnetic field (magnetotaxis). Although its ecological function is uncertain, magnetotaxis is assumed to play a role in determining the ideal oxygen concentrations. Magnetosomes are encased in a lipid bilayer membrane and made of the mineral magnetite or greigite. The morphology of magnetosomes is species-specific. Endopsores The development of endospores is arguably the most well-known bacterial response to stress. Endospores are bacterial survival structures that are extremely resilient to a variety of chemical and environmental challenges, allowing bacteria to survive in conditions that would be fatal to them in their usual vegetative state. It has been proposed that endospore creation has allowed for the survival of some bacteria for hundreds of millions of years (e.g. in salt crystals) although these articles have been questioned. Certain genera of gram-positive bacteria, including Bacillus and Clostridium, are the only ones that can produce endospores. Only one spore is produced per cell, unlike reproductive spores, hence there is no overall increase in the number of cells after endospore germination. An endospore's location within a cell can be utilized to identify a bacterium because it is species-specific. Dipicolinic acid is a substance that makes up 5% to 15% of the dry weight of bacterial spores and is thought to be the reason why endospores can withstand heat. Researchers have identified living endospores retrieved from the intestines of Egyptian mummies as well as from lake sediments in Northern Sweden estimated to be many thousands of years old. Cytoskeleton The prokaryotic cytoskeleton is the aggregate name for all structural filaments in prokaryotes. It was formerly considered that prokaryotic cells did not possess cytoskeletons. However, breakthroughs in imaging technology and structural determination have demonstrated the presence of filaments in these cells. Prokaryotes contain homologs for every significant cytoskeletal protein discovered in eukaryotes. Cytoskeletal components perform crucial functions in cell division, protection, shape formation, and polarity determination in diverse prokaryotes.
Next TopicSystolic Blood Pressure
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








