| |

Prokaryotic Cell DefinitionA prokaryotic cell misses the mark on core and other layer-bound organelles. Prokaryotes are unicellular creatures that comprise bacteria and archaea. The genetic material of prokaryotic cells is contained in a single circular chromosome found in the nucleoid area of the cell. Prokaryotic cells have a cell layer, cytoplasm, ribosomes, and a cell wall that offers immediate help and security. Certain prokaryotic cells may also contain flagella for movement and pili for surface attachment. 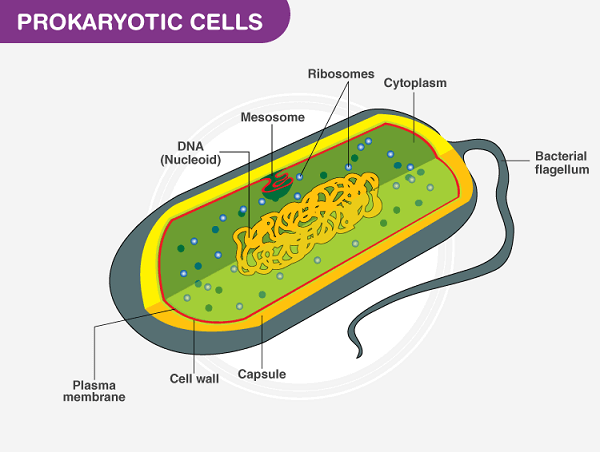
Prokaryote is derived from the Greek words (expert, 'previously') and (karyon, 'nut' or 'bit'). Prokaryotes were grouped into the realm of Prokaryota in �douard Chatton's two-domain paradigm. Nevertheless, under the three-space framework, prokaryotes are divided into two categories based on atomic examination: Microscopic Organisms (formerly Eubacteria) and Archaea (previously Archaebacteria). Eukaryota is the third space for life forms with cores. Prokaryotes are remembered to have emerged before eukaryotes in a natural turn of events. Apart from lacking a core, prokaryotes require mitochondria or the bulk of the other layer-bound organelles that characterize the eukaryotic cell. It was formerly thought that prokaryotic cellular elements inside the cytoplasm were unenclosed, save from an exterior cell film; however, bacterial microcompartments, thought to be fundamental organelles contained in protein shells, have been discovered alongside other prokaryotic organelles. Certain prokaryotes, like cyanobacteria, may create enormous provinces while being unicellular. Some have multicellular stages throughout their daily life cycles, such as myxobacteria. Prokaryotes are abiogenesis, reproducing without gametes; however, a flat-quality exchange may occur. Nuclear exploration has given knowledge into the advancement and reliance of the three spaces of life. The qualification among prokaryotes and eukaryotes mirrors the presence of two particular degrees of cellular association; just eukaryotic cells have an encased center that incorporates chromosomal DNA and other trademark layer-bound organelles, for example, mitochondria. Extremophiles and Methanogens are two distinct types of prokaryotes that are common in a variety of extreme environments. Structure of Prokaryotic CellProkaryotic cells have a straightforward construction coordinated around a focal nucleoid district. The nucleoid district is the site of DNA replication and recording, and it contains a solitary round chromosome that isn't encased by an atomic layer. The DNA in prokaryotic cells is coordinated into a minimal design called a nucleoid, which is moored to the cell layer. 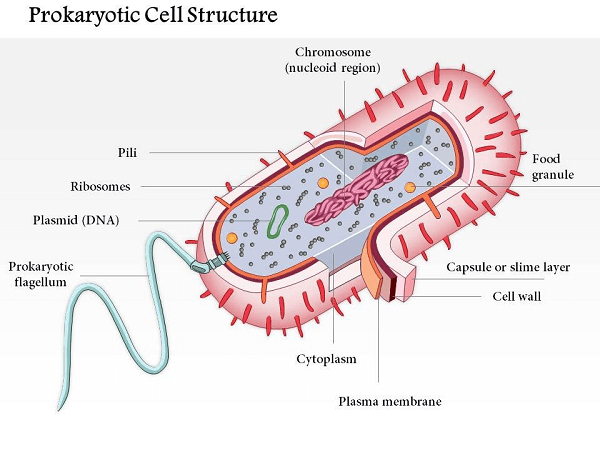
The cell layer of prokaryotic cells is a lipid bilayer that encases the cytoplasm and isolates the cell from its current circumstance. The cell layer is specifically porous and manages the development of materials into and out of the cell. The cytoplasm of prokaryotic cells contains the cell all's inner designs and organelles. A thick arrangement contains different macromolecules, including proteins, nucleic acids, and compounds. Ribosomes are the most bountiful organelles in the cytoplasm and are answerable for a protein blend. Prokaryotic cells have an unbending cell wall that offers immediate help and security. The cell wall is made of an exceptional particle called peptidoglycan, which isn't tracked down in eukaryotic cells. The cell wall additionally keeps up with the state of the cell and keeps it from blasting under high osmotic strain. A few prokaryotic cells have different outer designs, like flagella and pili. Flagella are long, whip-like designs utilized for headway, while pili are more limited, hair-like designs utilized for connection to surfaces. Metabolic Pathways of Prokaryotic cellProkaryotic cells have a different scope of metabolic pathways that permit them to get energy from various sources. They can be grouped into three general classifications: Photoautotrophs, Chemoautotrophs, and Heterotrophs. 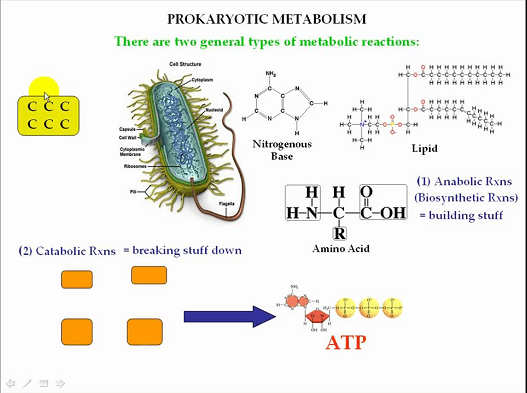
Photoautotrophs utilize light energy to deliver natural mixtures from carbon dioxide and water. They can photosynthesize utilizing shades like chlorophyll or bacteriochlorophyll. The most notable photoautotrophs are plants and green growth; however, numerous prokaryotic life forms, like cyanobacteria, additionally use photosynthesis. Chemoautotrophs acquire energy from inorganic atoms like smelling salts, nitrites, or sulfur. They can likewise involve carbon dioxide as their wellspring of carbon. Chemoautotrophs are found in outrageous conditions, for example, remote ocean aqueous vents, where there is no light, and the natural matter is scant. Heterotrophs acquire energy from natural atoms, like glucose, requiring an outer wellspring of carbon. Heterotrophs can be arranged into three subcategories given their energy source: high-impact, anaerobic, and facultatively anaerobic. DNA TransferEven though it has been mostly studied in microorganisms, DNA movement across prokaryotic cells occurs in archaea and bacteria. Three rounds of quality exchange occur in microorganisms. They are bacterial infection (bacteriophage)-intervened transduction, plasmid-intervened creation, and regular change. Bacteriophage transduction of bacterial properties appears to be a rare mishap during intracellular collecting of infection particles rather than a mutation of the host microscopic organisms. The exchange of bacterial DNA is primarily affected by bacteriophage properties rather than bacterial traits. It is a transformation for disseminating copies of a plasmid, starting with one bacterial host and then onto the next. It is controlled by plasmid attributes in the highly focused E. coli framework. A plasmid may randomly integrate into the host bacterial chromosome during this cycle, transferring a portion of the host bacterial DNA to another bacterium. A bacterial transformation is not immediately apparent when a plasmid intervenes in the movement of host bacterial DNA (formation). 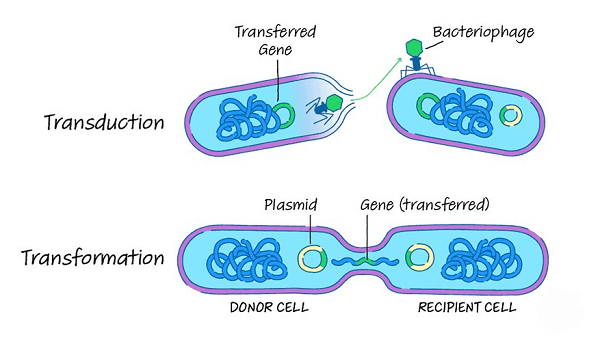
The normal bacterial change includes the movement of DNA from one bacterium to the next via the interceding media. Unlike transduction and creation, change is a bacterial transformation for DNA movement since it relies on multiple bacterial quality items that expressly collaborate to carry out this intricate cycle. A bacteria must first enter an exceptional physiological condition known as a skill to bind, pick up, and recombine donor DNA into its chromosome. Bacillus subtilis is believed to have around 40 attributes that will boost capability. The length of DNA transferred during B. subtilis transformation can be as much as a third of the length of the complete chromosome. Change is a common technique for DNA movement, and 67 prokaryotic species are known to be ordinarily competent for change. 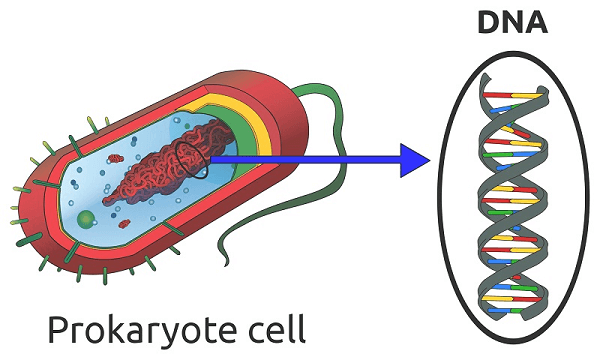
Among archaea, Halobacterium volcanii structures cytoplasmic frameworks between cells that appear to be used to move DNA, beginning with one cell and then onto the following. Another archaeon, Sulfolobus solfataricus, moves DNA between cells by direct contact. Relationship to EukaryotesThe division between prokaryotes and eukaryotes is typically considered the main qualification or distinction among living beings. The differentiation is that eukaryotic cells have a "valid" core containing their DNA, while prokaryotic cells don't have a core. Although both eukaryotes and prokaryotes have massive RNA/protein complexes called ribosomes that make protein, the ribosomes of prokaryotes are more modest than those of eukaryotes. Mitochondria and chloroplasts, two organelles seen in numerous eukaryotic cells, incorporate ribosomes similar in size and appearance to those tracked down in prokaryotes. This is one of a few bits of proof showing mitochondria and chloroplasts are vanishing from free-living microscopic organisms. According to the endosymbiotic concept, early eukaryotic cells phagocytosed primitive prokaryotic cells and modified them to solidify their designs, resulting in mitochondria and chloroplasts. A prokaryote's genome is contained within the nucleoid, a DNA/protein complex in the cytoplasm that falls short of the atomic envelope. In contrast to the several direct, conservative, excellently coordinated chromosomes discovered in eukaryotic cells, the complex includes a single, cyclic, doubly abandoned particle of stable chromosomal DNA. Moreover, many important properties of prokaryotes are stored in distinct circular DNA structures known as plasmids. Prokaryotes, like Eukaryotes, may replicate hereditary material and have a haploid chromosomal structure that is partially repetitive, a situation known as merodiploid. 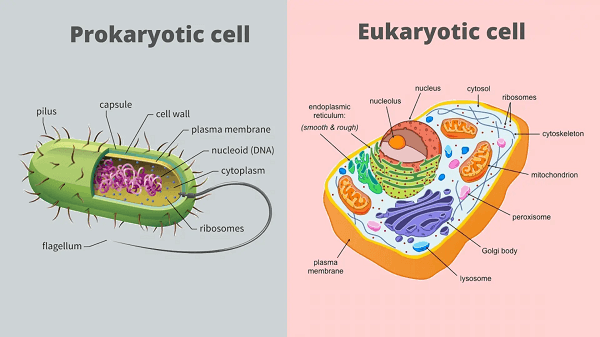
Prokaryotes need mitochondria and chloroplasts. Cycles, for example, oxidative phosphorylation and photosynthesis, happen across the prokaryotic cell layer. Be that as it may, prokaryotes, in all actuality, do have a few inside structures, like prokaryotic cytoskeletons. It has been suggested that the bacterial phylum Planctomycetota has a layer around the nucleoid and contains other film-bound cellular plans. Further assessment uncovered that Planctomycetota cells are interconnected and not categorized or nucleated as other bacterial film structures. Prokaryotic cells are typically a lot more modest than eukaryotic cells. Thusly, prokaryotes have a bigger surface-region-to-volume proportion, giving them a higher metabolic rate, a higher development rate, and as an outcome, a more limited age time than eukaryotes. There is mounting evidence that the eukaryotic foundations are to be found in (or near to) the archaea asgard family, perhaps Heimdallarchaeota (a thought which is a cutting edge variant of the 1984 eucyte speculation, eucyte being an old equivalent for Thermoproteota, a taxon to be found close by the then-obscure Asgard bunch) Histones, which generally bundle DNA in eukaryotic cores, have been found in a few archaean assemblages, demonstrating homology. According to the endosymbiotic concept, this odd ancestor of eukaryotic cells (eucytes) overwhelmed an Alphaproteobacteria, creating the first eucyte (LECA, the last eukaryotic normal precursor). There could have been some extra help from infections, called viral eukaryogenesis. ThomasArrogant Smith named the non-bacterial assemblage of archaea and eukaryota Neomura in 2002. But, from a cladistic perspective, since eukaryotes evolved from the group of dinosaurs known as maniraptorans, they are archaea similar to how birds are dinosaurs. Conversely, archaea without eukaryota give off an impression of being a paraphyletic bunch, much like dinosaurs without birds. Cell SizeAverage prokaryotic cells range from 0.1 to 5.0 micrometers (?m) in breadth and are more modest than eukaryotic cells, which have widths from 10 to 100 ?m. The figure underneath shows the spans of prokaryotic, bacterial, and eukaryotic, plant and creature cells and different particles and organic entities on a logarithmic scale. Every unit of expansion in a logarithmic scale addresses a 10-crease expansion in the estimated amount, so we're discussing these enormous size contrasts! 
Except for a few interesting exceptions, such as the single-celled kelp Caulerpa, cells should stay microscopic, whether prokaryotic or eukaryotic. What is the explanation behind this situation? The key answer is that as cells grow, it becomes increasingly difficult to trade an acceptable amount of nutrients and waste with their present situation. To understand how this works, consider the surface-region-to-volume ratio of a cell. The surface-region-to-volume ratio is crucial because the plasma film is the cell's interface with the environment. If the cell wants to take up vitamins, it should do so across the film, and if it has to remove waste, the layer is once again its only channel. Since it has a fixed amount of channels, each layer fix can only exchange so much of a specific material in a given interval. If the cell grows too large, its layer will not have enough trade limit (surface region, square capacity) to aid the rate of trade predicted for its enlarged metabolic movement. The surface-area-to-volume problem is just one of several issues brought on by large cell sizes. Moving materials within cells take longer as they get larger. These considerations set an overall maximum breaking point for cell size, with eukaryotic cells potentially outnumbering prokaryotic cells due to their main and metabolic components. A few cells likewise utilize mathematical stunts to get around the surface-region-to-volume issue. For example, a few cells are long and thin or have numerous bulges from their surface, including that increment surface region compared with volume squared. Characteristics of Prokaryotic CellAll prokaryotic cells have a nucleoid region, DNA and RNA serving as the genetic building blocks, ribosomes producing proteins, and cytoplasm containing a cytoskeleton organizing the cellular components. Yet, prokaryotic organisms are a hugely diverse collection of species that come in a wide variety of shapes and sizes. These structural changes often mirror functional changes, and these many distinct creatures inhabit varied habitats. Prokaryotic cells are typically between 0.1 and 5 micrometers long (.00001 to .0005 cm). Eukaryotic cells are substantially bigger, measuring between 10 and 100 micrometers. Because prokaryotic cells are smaller, they have a greater surface-area-to-volume ratio, allowing them to acquire more nutrients via their plasma membrane. How Do Prokaryotic Cells DivideBinary Fission is the interaction through which prokaryotic cells partition. Dissimilar to mitosis, this component does exclude DNA buildup or organelle duplication. 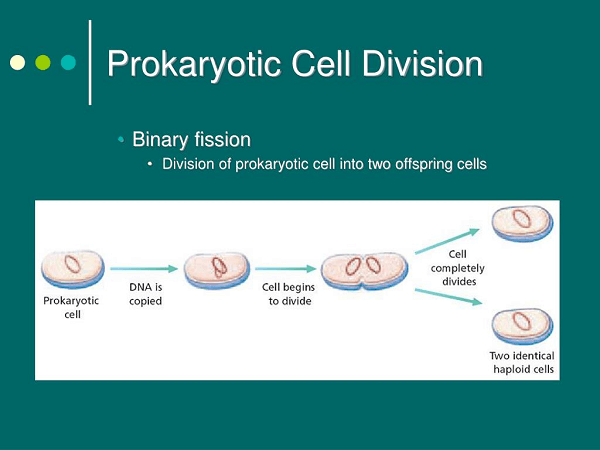
Prokaryotic cells have no DNA, which isn't put away in complex chromosomes. In addition, since there are no organelles, nothing remains to be parted. As a prokaryote creates an incredible size, it goes through double splitting. This method duplicates the DNA and separates each new strand into unmistakable cells. This is a less complex interaction than mitosis. Microscopic organisms can increase essentially faster than most eukaryotic animals.
Next TopicTrapezium Definition
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








