| |

AuxinThe discovery of auxins was the beginning of the primary plant hormones studies. Their name comes from the Greek word v (auxein, which means "to grow/increase"). Auxin can be found in all regions of a plant, albeit in varying amounts. Because the concentration in each site contains important developmental information, it is tightly regulated by metabolism and transport. As a result, auxin produces "patterns" of auxin intensity maxima and minima in the plant organs, which regulate the development of individual cells and, eventually, the entire plant. 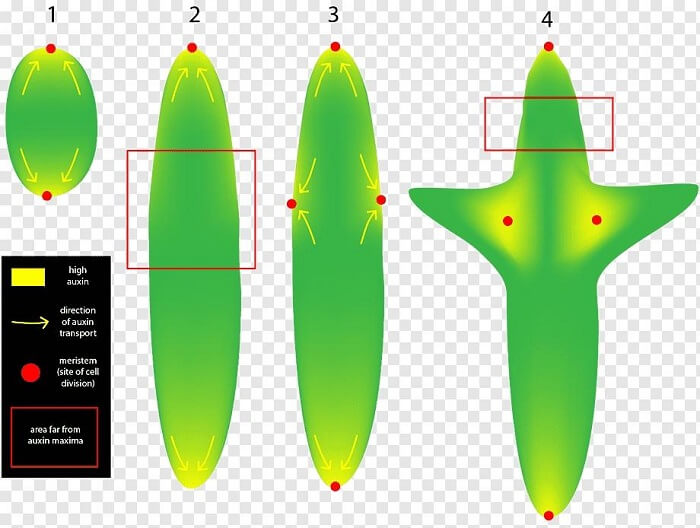
The dynamic and environment-responsive structure of auxin distribution inside the plant is a critical component in plant growth, environmental response, and, specifically, plant organ development (such as leaves or flowers). It is accomplished through the so-called polar auxin transport, which is a highly complicated and well-coordinated active movement of auxin molecules from cell to cell all across the plant body. As a result, a plant can react to and adjust to environmental conditions (as a whole) without the need for a nervous system. Auxins usually work in tandem with other plant hormones, or in conflict to them. The proportion of auxin to cytokinin in some plant tissues, for example, controls whether root or shoot buds form. What is Auxin?Auxins are a series of plant hormones that can be found naturally or synthesized artificially. They serve a crucial function in plant growth regulation. Auxins were first discovered in human urine. The word auxin means "enlarge" or "increase." Cell division, differentiation, and elongation are all induced by them. Auxin(s) could be defined as a group of chemicals that influence plant growth in the same way that IAA does. Other natural auxins have been discovered in plants, including 4-Cl-IAA and PAA, while some synthetic chemicals, such as NAA or 2, 4-D, have IAA-like function and are widely employed in horticulture, farming, and research. HistoryThe most researched auxin, IAA, was discovered in maize by chemists in the early 1900s, but its prevalence had been suspected for decades. In the petiole of canary grass, Charles Darwin noticed phototropism movement (plants bending towards light). He saw that near the tip of the coleoptile, there was an influencer that caused the curving towards the light. The chemical involved for phototropic movement in the oat coleoptile was later isolated and termed "Auxin" by Frits Went. Kenneth Thimann refined and deciphered the structures of main auxins such as IAA (Indoleacetic acid). These early discoveries sparked the growth of a vibrant and active study sector that has yielded significant results in recent decades. Auxin regulates cell growth at the meristems, which determine plant architecture, and appears to affect practically all developmental phases in plants, from early embryogenesis through fruit ripening. The goal of current research is to figure out how such a tiny molecule may be ubiquitous while also having context-dependent function. Type of AuxinsNaturally Occurring Auxins: Indole-3-acetic acid (IAA), Indole butyric acid (IBA). Artificially synthesized Auxins: 2,4-dichloro phenoxy acetic acid (2,4-D), Naphthalene acetic acid (NAA) Hormonal activityAuxins aid plant development at all stages, from the molecular level to the organ level and finally to the entire plant. Molecular mechanismsWhen auxin interacts with a plant cell, it causes substantial changes in gene expression patterns, with many genes being up- or down-regulated. The particular methods by which this happens are still being investigated, but at least 2 auxin signaling routes have been identified. PerceptionThe TIR1/ AFB class of F-box proteins are the most well-studied auxin receptors. F-box proteins use the ubiquitin disintegration pathway to engage other proteins for degradation. When TIR1/ AFB protein connect to auxin, the auxin functions as a 'molecular glue,' allowing them to bind to their targets. ABP1 is another auxin-binding protein that is now commonly accepted as an auxin receptors (at the apoplast). However it is thought to play a considerably smaller impact than the TIR1/AFB signaling pathway, and little is known about ABP1 signaling. Aux/IAA and ARF signaling modulesAuxin signaling- When auxin is absent, Aux/IAA bind to ARFs and decrease their transcriptional activity. When auxin is present, it produces a "molecular glue" among TIR1 and Aux/IAAs, causing these repressors to degrade. ARFs are thus free to attach to DNA and produce transcriptional modifications. Auxin responsive factors (ARFs) are a class of regulatory factors that play a role in auxin signaling. ARFs engage to a class of repressors known as Aux/IAAs in the absence of auxin. Aux/IAA inhibit ARFs' ability to increase gene transcription. Furthermore, Aux/IAA binds to ARFs, bringing it into interaction with the regulators of auxin-regulated genes. When Aux/IAA binds to these promoters, it represses gene expression by enlisting the help of other factors to alter the DNA structure. Auxin binds to TIR1/AFBs, allowing them to connect to Aux/IAAs. Aux/IAAs are indicated for degradation when bound by TIR1/AFBs. When Aux/IAA is degraded, ARF proteins are released, allowing them to stimulate or repress genes whose terminals they are linked to. The astounding diversity of auxin responses is assumed to be due to the huge amount of Aux/IAA and ARF binding combinations that are available, as well as their varied distributions throughout cell types and developmental stages. On a cellular levelAuxin is required for cell growth at the cellular level, impacting both cell division and cellular growth. Auxin concentration, in combination with other local variables, plays a role in cell differentiation and destiny specification. Auxin can stimulate axial lengthening (as in shoots), flexural capacity (as in root dilatation), or iso-diametric expansion, depending on the tissue (as in fruit growth). Auxin-promoted cellular expansion happens in the absence of cell division in some situations (coleoptile growth). Auxin-promoted cell division and cell growth may occur in close succession within the same tissue in other instances (root initiation, fruit growth). Auxins and other plant hormones almost always appear to interact in a living plant to influence plant development patterns. Organ patternsPlant tissue grows as a result of the growth and division of plant cells, and particular tissue growth helps in the evolution of plant organs. Unevenly localized growth causes organs to bend, turn, and directionalize-for instance, stems spinning toward light sources (phototropism), roots development in reaction to gravity (gravitropism), and other tropisms emerged because cells on one side of the organ grow faster than cells on the other side of the organ. As a result, careful assessment of auxin distribution amongst distinct cells is critical to the plants overall growth and organization. Organization of the plantAuxins are essential for the healthy establishment of the plant since they contribute to organ shape. Plants would just proliferate heaps of similar cells if hormonal regulation and organization were absent. Auxin deployment begins in the plant embryo, where directional auxin distribution promotes subsequent advancement of main growth poles, which leads to the formation of future organ buds. It also aids in the appropriate development of emerging organs such as roots, cotyledons, and leaves, as well as transmits long-distance signals between them, adding to the plant's overall architecture. Throughout the plant's life, auxin aids in maintaining polarity of growth and allowing the plant to "recognize" where its branches (or any organ) are connected. Mechanism of Action of Auxin
Next TopicGram Positive Bacteria
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








