| |

Bacteria Cell SizeOur perception of microorganisms is largely affected by their small size. The existence of most of these ancient species was not understood until the 17th century, when the microscope was invented. Initially thought to be little more than "bags of enzymes," subsequent discoveries in imaging, molecular phylogeny, and, most recently, genomes have shown remarkable variety inside this hitherto unseen world of life. The influence that bacteria have on its evolution, physiology, and morphogenesis is discussed in this article. 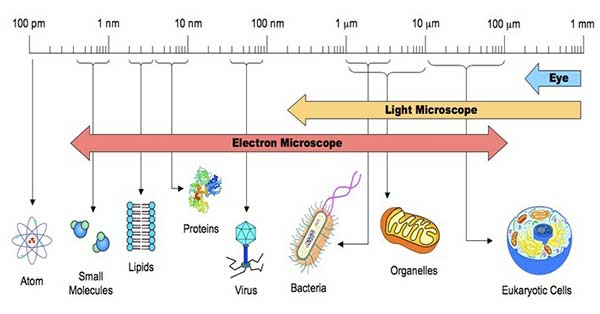
CandidatusActinomarinaminuta is a free-living marine ultramicrobacteria with a cell volume that is 1 percent that of E. coli (0.013 mm3, range 0.6 1022 to 2.4 1022 fL). The marine sediment-dwelling bacteria, often known as "Sulfur pearl of Namibia," is a spherical bacteria with eight times larger than E. coli (750 mm in diameter, volume 2.2 108 fL or 0.22 mL). The scientific name of this bacteria is Thiomargaritanamibiensis. Thiomargarita is somewhat bigger than a Drosophila eye and large enough to be visible by the human eye. A closer examination of Thiomargarita has a centrally positioned fluid-filled vacuole that occupies 98 percent of the cell volume and serves as a nitrate reservoir for sulfide oxidation. Even when the internal vacuole is considered, a giant Thiomargarita cell has a huge biovolume to sustain (4.4 106 fL). Epulopiscium spp., the biggest known heterotrophic bacteria, are intestinal symbionts of some marine surgeonfish. These cigar-shaped cells have an active cytoplasmic capacity of 2 106 mm3 (0.02 mL) and a diameter of up to 600 mm and 80 mm. Epulopiscium cells, unlike Thiomargarita, lack storage vacuoles and other inert inclusions. CandidatusActinomarina's minute size can be compared to that of behemoths such that the gap between both the species is just like a mouse and the Empire State Building (E. coli could be represented by a small skunk or a rabbit on this scale). 
What Factors Influence the Size of Bacterial Cells?The tiniest cells need sufficient room to contain sufficient genetic resources to maintain their lifestyle. To sustain its metabolism and cellular reproduction, the cell must also include the fundamental machinery necessary to express those genes and housekeeping proteins and biochemicals. Obligate intracellular symbionts, pathogens, and organelles that have given up metabolic capacities because their demands are satisfied by the host exhibit genomic and metabolic streamlining. The loss of genes involved in perceiving and reacting to environmental changes may result in significant genome reduction, but not necessarily in a proportional decrease in cell size. The shape and function of all big cells seem to be constrained by diffusion's constraints. The capacity of big bacteria to survive in their environment is influenced by interactions with nutrients, waste removal, and the timely movement of biomolecules inside the cell to meet metabolic demands. Eukaryotic cells are more advanced than bacterial cells, and they have a comparatively large size. This advancement and size credit goes to the compartmentalization of cellular functions in the Eukaryotic cells. Not only this, but they also contain cytoskeleton network protein trafficking along with the expansion of genomic resources. They have also acquired symbiotic organisms, which are energy-generating organelles. The Diffusion ProblemThe discovery of enormous bacteria necessitated a rethinking of long-held views regarding bacterial cell size limits. Diffusion is a molecule's random three-dimensional movement. It is the most effective method of transporting molecules inside a cell across a small distance of a few microns. Even a big protein may dependably travel this distance in less than a second in the cytoplasm. As a result, a biomolecule produced or introduced into a normal bacterium has a high possibility of reaching its active site practically rapidly. However, when measured on our scale, diffusive transport over a millimeter gap-roughly the length of an Epulopiscium cell and a fraction of the circle of Thiomargarita-becomes numbingly inaccurate. At ambient temperature, a tiny molecule like oxygen would take around one hour to diffuse 1 mm (Schulz and Jorgensen 2001). Similarly, nutrient uptake is dependent on the diffusion and capture of molecules at a cell's surface. As a result, free-living cells are tiny, with a high surface area compared to their cytoplasmic volume, allowing for the collection of nutrients at low concentrations to meet the metabolic demands of the cell. These restrictions have been placed on cell shape and function. E. coli has a surface-area-to-volume ratio of 3.7 mm2 to 1 mm3, whereas the biggest Thiomargarita cell had a surface-area-to-volume ratio of 8.2 1023 mm2 to 1 mm3. These enormous germs are clearly breaking the rules. The Reproduction of The GiantsEnormous bacteria have a variety of reproductive techniques, some of which may aid in maximizing their reproductive potential, development, and release of large progeny. All of the Epulopiscium spp. that are the biggest reproduce once a day, producing two or more intracellular progeny. Surprisingly, the commencement and development of internal offspring follow a regular daily cycle, and any particular population inside a host fish is highly coordinated in terms of development. The development of the offspring happens throughout the day while the host is eating. The mother cell divides bipolarly to start the process of reproduction. The polar cells are entirely swallowed, and the children develop within a membrane-bound compartment in the cytoplasm of the mother cell until it is completely filled. Offspring emerge through a split in the mother cell envelope in the final stages. Despite their developmental synchronization, the volume of a single population may vary by a factor of five. The mother cell seems to undergo a sort of programmed cell death at a late stage of development, a procedure that presumably conserves biochemical resources collected throughout growth. Epulopiscium was first categorized as a new protist due to its size, atypical ways of reproduction, and the difficulties in recognizing evolutionary connections among bacteria based on morphological traits. Endospore development led to internal progeny generation in Epulopiscium spp. The spore-forming Firmicutes seem to have adapted endospore creation as a reproduction mechanism multiple times. 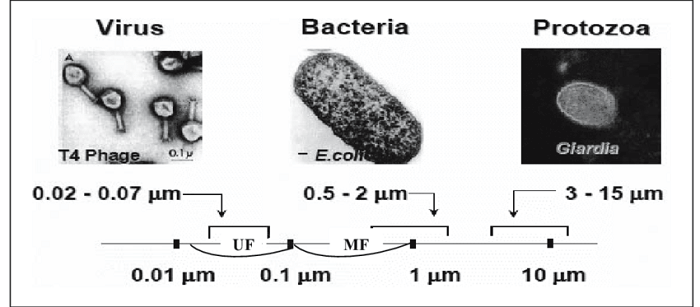
The Impact of The Environment on The Size of SpeciesLarge bacteria thrive in conditions where resources are accessible on a constant basis and concentrations are high. These correlations provide some insight into the circumstances that may enable organisms to overcome cell size diffusion constraints. Giant spirochetes may be found in nutrient-rich sediments and various digestive systems, including termites' hindgut. Several herbivores' digestive tracts have been shown to have large rod-shaped cells. Thiomargarita, a sulfur-oxidizing bacterial cell, is abundant in marine sediments. They are found almost everywhere on the planet and take minerals with them to power their breathing. A plentiful source of energy is a recurring character in all of these systems, and it may serve as a unifying factor. Polyploidy is common in bacteria, although it is costly to maintain. Selection for scaling down to a single copy of the genome may be eased in nutrient-rich settings or when energy is nearly inexhaustible, allowing polyploid bacteria to flourish. Following that, changes to the cellular design may be made to allow for even further size increase. 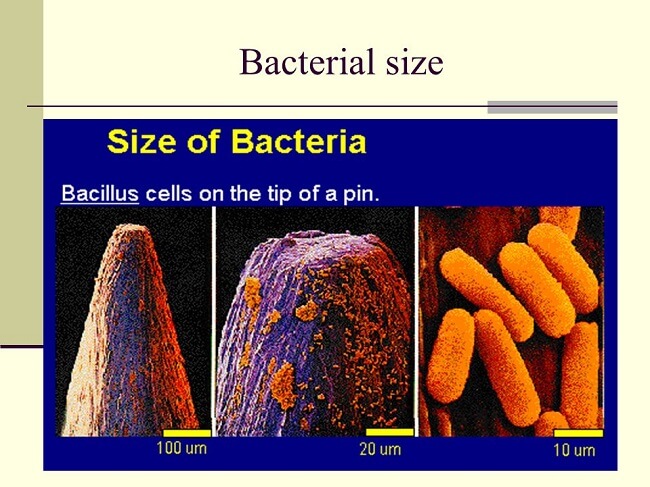
Homeostasis Of Cell SizeIsogenic populations of bacteria retain cell size within very tight boundaries under steady-state settings. For each division event, bacteria commence and end one cycle of DNA replication, similar to eukaryotes. In contrast to eukaryotes, DNA replication, chromosomal segregation, and assembly of the division machinery are not separate operations but rather overlap to varying degrees depending on growth pace. Importantly, bacteria can sustain inter-division durations less than half the amount of time necessary for chromosomal DNA replication under rapid growth rates maintained by nutrient-rich circumstances. E. coli, for example, may double in size and split in as little as 20 minutes; nevertheless, replication of the complete 4-Mb E. coli chromosome takes 60 minutes even under optimal circumstances. Rapidly expanding cells seem to overcome this apparent contradiction by continuous manufacturing of DNA and launching new replication rounds before the old one is completed. These cells may have 4, 8, or even 16 replication forks active at any one time. Despite the fact that initiation of replication is limited to only once during cell cycle division, only one round of replication is required before division. Thus, it can be said that the initiation event and termination events are not always co-related with the same replication fork within a single division cycle. Controlling cell division and cell size in most bacteria division begins forming a ring-like structure at the prospective division site by the crucial GTPaseFtsZ. The "Z-ring" acts as scaffolding for the remainder of the cell division machinery to be assembled. Although the two proteins have little sequence resemblance, structural research shows that FtsZ is an evolutionary predecessor of tubulin. At any one moment, around one-third of the FtsZ in the cell is in the FtsZ ring, with subunit turnover rates on the order of seconds. The cytoskeletal protein FtsZ directly influences cell size, making it a suitable target for variables influencing cell size homeostasis. FtsZ depletion has little impact on cell growth or DNA replication at first but causes a fast rise in length. Under steady-state circumstances, even small decreases (20%) in intracellular FtsZ concentration result in a substantial increase (.50%) in cell length. It's simple to envision how variations in the supply of bricks would affect cell size if the cytokinetic ring were a brick construction. Bricks are easily accessible when FtsZ concentration is high, decreasing travel time and enabling the construction of a stable and functioning Z-ring. Reduced FtsZ availability, on the other hand, slows the delivery of bricks to the building site, delaying the assembly of the division equipment and, with it, cytokinesis. Delays in the construction of the cytokinetic ring result in increases in cell size because cells continue to grow at normal rates. In E. coli and B. subtilis, the levels of FtsZ are directly proportional to the size of the cell, and it is independent of the growth rate. Stochastic changes in cell size should be greatly reduced by FtsZ availability. Short cells will take longer to gather enough FtsZ to divide, resulting in a larger division size. On the other hand, longer cells will need less time to collect enough FtsZ to facilitate division machinery assembly, resulting in shorter interdivision times and shorter daughter-cell length. Only the accumulation of FtsZ alone is not adequate to begin cell division in bacteria. It is the same as DnaA, whose accumulation itself is not enough to account for cell size homeostasis. Despite the fact that two-fold overexpression of FtsZ lowers cell size by 10% in both E. coli and B. subtilis, it has no effect on the time of division. It's yet unclear how these variables interact with cell-cycle progression. FtsZ is an important component of cytokinetic cell machinery. It is involved in the rate-limiting step of cell division, acting like a potential candidate for cell homeostatic regulation. Cell Size Regulation Depends on NutrientsIn many bacteria, nutrient availability is the major factor in cell size. Schaechter, Maale, and Kjeldgaard discovered that Salmonella cell size changes by up to twice depending on growth rate in a classic article. The size of cells cultivated in nutrient-rich media with doubling periods of 20 minutes was more than double that of cells cultured in nutrient-poor medium with mass doubling durations of well over an hour. Regardless of any growth parameter or if the growth is delayed due to any other reason, the link between growth rate and size remains constant. Even if the bacteria is getting through some stress or there is any delay in carbon, nitrogen, and phosphate nutrition, the cell size is not affected. This suggests that the fundamental driver of the size of the cell is the doubling time and not the content of nutrients it is getting in the medium. However, recent research reveals that bacterial size is a complicated process involving a well-choreographed series of regulatory circuits, each of which is receptive to a separate and sometimes overlapping set of nutrient-dependent signals.
Next TopicBacterial Shapes
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








