| |

Basidiomycete, Basidium, And BasidiosporesBasidiomycota refers to one of two main divisions that make up the subkingdom Dikarya (also known as higher fungi) within the fungi Kingdom, along with Ascomycota. So, Basidiomycetes are the members. Basidiomycetes have distinct club-shaped bodies, which are the fruiting bodies of these fungi. These fruiting bodies are called basidia (singular, basidium). These basidia will be visible as distinct structures if we observe basidiomycetes under the light microscope. 
These basidia are the main reproductive bodies found in all species of basidiomycetes. The most common examples of this fungus are mushrooms. They can be poisonous or eatable. They are commonly found in moist and warm places growing on the wood bark or land. Mushrooms can be easily found in marketplaces and can be just growing on your lawn. A basidiomycete's fruiting bodies create a ring in a meadow, commonly referred to as a "fairy ring." The scientific name for the most well-known fairy ring fungus is Marasmius oreades. This fungus's body, mycelium, is underground and grows in a circle. The role of mycelium is to take the nitrogen from the soil as the fungi develop. This results in the spreading of mycelia from the centre in various locations, thus forming a fairy ring. A fairy ring of fruiting bodies is formed where enough nitrogen is present to sustain them. The mushroom body growing outside the soil has a cap-like structure. This cap has gills on the underside; therefore, this mushroom-producing fungus is also called gill fungus. The "gills" are compressed hyphae that carry the basidia. Shelf fungus, which clings to the bark of trees like little shelves, is also included in this category. The Basidiomycota also includes smuts and rusts, which are major plant diseases, as well as toadstools. The Phylum Basidiomycota contains the majority of edible fungus, yet, certain basidiomycetes release lethal poisons. Cryptococcus neoformans, for example, cause severe respiratory disease. In the life cycle of basidiomycetes, both asexual and sexual reproduction is present. The other fungi are in the haplontic stage for most of their lives. But basidiomycete is an advanced group, producing both haplontic and dikaryotic mycelium. (Technically, the dikaryotic phase is not diploid since the nuclei remain unfused until soon before spore generation.) 
Sexual spores are more abundant in basidiomycetes than asexual spores. Basidiospores are sexual spores that develop in the club-shaped basidium. The nuclei of two distinct mating strains combine (karyogamy) in the basidium, producing a diploid zygote that undergoes meiosis. The haploid nuclei travel into four chambers attached to the basidium, where they mature into basidiospores. Each basidiospore germinates and produces haploid monokaryotic hyphae. The resulting mycelium is known as the main mycelium. Mycelia from distinct mating strains can unite to form a secondary mycelium with haploid nuclei from both mating strains. This dikaryotic stage dominates the basidiomycete life cycle. As a result, each cell in this mycelium contains two haploid nuclei that will not fuse until the basidium forms. It is the secondary mycelium which is responsible for producing the basidiocarp. Basidiocarp is a fruiting body that comes out of the ground in the form of a mushroom. The growing basidia are seen on the gills beneath the cap of the basidiocarp. BasidiocarpIn fungi, spores are responsible for the production of the body. Spores are produced within a huge sporophore (fruiting body). Spores are produced via sexual reproduction inside a sporophore, which is grown on the surface of a club-shaped structure called basidia. The fruiting body is called basidiocarp. In the phylum Basidiomycota, all the members produce basidiocarp except smut and rust fungi. 
The biggest basidiocarps include gigantic puffballs (Calvatia gigantea), which may grow to be 1.6 m (5.25 feet) long, 1.35 m wide, and 24 cm (9.5 inches) tall, and bracket fungus (Polyporus squamosus), which can grow to be 2 m in diameter. Single cells of the yeast-like Sporobolomyces are the tiniest. The mature meiosporangium (basidium) structure appears to be evolutionarily conservative; hence, physical similarities between significant taxa reflect varying degrees of connection between such taxa. The basidium is the organ where karyogamy and meiosis occur and where meiospores (basidiospores) form. Because fossils of basidiomycete basidiocarps are either few or difficult to interpret, comparative morphological studies appear to be the only viable way of studying evolution within the Basidiomycotina. The classes Heterobasidiomycetes and Homobasidiomycetes are the two primary taxa of the subdivision Basidiomycotina. Heterobasidiomycetes are organisms that produce basidiospores capable of generating secondary spores and/or yeast-like cells. Homobasidiomycetes are taxa that cannot generate secondary spores and/or yeast-like cells. Basidium
The basidium is a club-like structure with 2-4 basidiospore-bearing apical sterigmata. It is normally whole, although Auriculariales and Platygloeales are split longitudinally. Tertiary mycelium is developed from secondary mycelium and is called hymenophore. Small fruiting bodies (spore-producing structures) are produced on hymenophores. The secondary mycelium is highly coiled and results in the formation of tertiary mycelium. It is a dikaryon, and one of the most distinguishing characteristics of basidiomycetes is basidia. A basidium typically produces four sexual spores known as basidiospores. However, this number can vary from two to eight. Each basidiospore in a typical basidium is carried at the tip of a slender prong, or horn called a sterigma (pl. sterigmata) and is violently released at maturity. The term basidium means "small pedestal," referring to how the basidium supports the spores. However, other scientists believe the structure is more like a club. Basidiole refers to an immature basidium. 
ArrangementMost Basidiomycota have single-celled basidia (holobasidia). However, other groups contain multicellular basidia (a phragmobasidia). PhragmobasidiumA phragmobasidium is a basidium that has been separated into distinct cells after meiosis, as in the Auriculariales and Platygloeales. The metabasidium (the area where meiosis occurs) appears as four cells in a cross-section. Sterigma (pl. Sterigmata)Sterigmata is an extension from the apical region of basidium that has basidiospores. In some basidiomycetes like Auriculariales and Platygloeales, basidium is not spherical but elongated. Each basidium has four sterigmata growing from its surface. CystidiaCystidia are sterile structures in certain Agaricales and Atheliales extending beyond the hymenium and occasionally coated with calcium oxalate crystals around their apices. In the Agaricales, distinct forms of cystidia can be found in different regions of the basidiocarp. The Basidium's Meiotic DivisionsThe reduction and recombination of chromosomal material is the primary purpose of meiotic divisions. Genetic studies suggest that these processes occur in a pretty typical manner in higher fungi. Nuclear division in higher fungi differs from the more well-known nuclear division in higher plants in several ways. 
Because of the consistent orientation of the division spindles, meiotic basidia of Homobasidiomycetes are ideal for studying nuclear division. In contrast to ascomycetes, basidiomycete's nuclear divisions have polar gaps or fenestrae throughout late anaphase and telophase. Biochemical and Genetic Investigations into the First Events of Fruitbody FormationFruit body development in higher basidiomycetes requires the expression of several structural and regulatory genes essential for morphogenetic processes. Under some environmental conditions, a genetic component may trigger the initial response required to commence fruiting. 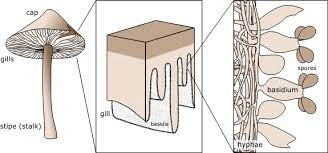
The incompatibility factors have been intensively explored since it has been demonstrated that they govern dikaryotization, which is required for fruiting in the normal growth process. Fruiting is not confined to dikaryotic mycelia; monokaryotic mycelia form fruitbodies under specific situations such as age, injury, or the impact of certain chemicals. Mutants that develop fruitbodies on monokaryotic mycelia have also been discovered. The study of monokaryotic fruiting benefits working with a basic system of fruiting initiation devoid of incompatibility considerations. Interactions between distinct sections of a mycelium and different mycelia are crucial in fungal morphogenesis and growth. The interactions that are better understood are mediated by chemicals released into the substrate and result in the commencement and development of sexual organs. Much less is known about interactions between distinct areas of individual hyphae or hyphal associations than regarding the evidence for the morphogenetic significance of secreted chemicals. BasidiosporesBasidiospores are reproductive spores generated by Basidiomycete fungus, including mushrooms, shelf fungi, rusts, and smuts. Basidiospores are formed by basidia, specialised fungal cells containing one haploid nucleus produced by meiosis. Four basidiospores grow on appendages from each basidium, two of which are of one strain and the other two of the opposing strain. Millions of basidia can be seen in the gills under the cap of one common species. 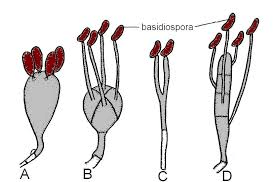
Some gilled mushrooms in the order of Agaricales can produce billions of spores. Calvatia gigantea, a puffball fungus, is estimated to produce five trillion basidiospores. Because most basidiospores are forcedly released, they are classified as ballistospores. These spores are the primary means of air dissemination for the fungus. The spores are discharged during periods of high humidity and have a peak concentration in the atmosphere at night or before sunrise. When basidiospores come into contact with a suitable substrate, they can germinate, often by generating hyphae. These hyphae spread from the initial spore to produce an expanding circle of mycelium. The round shape of a fungal colony explains how fairy rings grow and the circular lesions caused by a skin-infecting fungus that causes ringworm. Some basidiospores reproduce by generating tiny spores instead of hyphae. Basidiospores develop on the sterigma of each basidium cell. At maturity, the spores are expelled a few millimetres from the sterigma. Basidiospores are 7.6 6 m long, hyaline, and oval. Once they have enough moisture, they develop fully and get discharged in 20 to 25 minutes. But there are some teliospores which may take time to germinate. They sometimes take upto 3 to 6 days or maybe more to germinate. Alternate soaking, drying, freezing, and thawing may aid in teliospore germination. The basidiospores are wind-borne to the alternative host, Berberis and Mahonia species, but seldom induce infection more than 180 to 270 m from the source. Basidiospores have a brief life cycle, lasting only a few hours under optimal circumstances. During basidiospores' germination, a germ tube is formed with a terminal appressorium. From this, an infection peg penetrates the epidermal cells directly. It is known that the fungus majorly infects the top surface of the leaf. Other parts of the plant, like berries and stems, also suffer some infections. Around 212 weeks after the leaf bud opens, vulnerable cultivars' leaves typically appear to develop resistance to penetration. Basidiospore Structure And ShapeBasidiospores are distinguished by the presence of an attachment peg (known as a hilar appendage) on their surface. The spore was connected to the basidium at this point. The hilar appendage is present in some of the Basidiomycetes, whereas it is absent in other basidiomycetes. 
Basidiospores are usually single-celled (no septa) and can be spherical, oval, oblong, ellipsoid, or cylindrical. The hilar appendage is prominent in certain basidiospores but not in others. An apical germ pore is also present in some basidiospores and is responsible for the asymmetric shape of the spore. The spore's surface might be relatively smooth or decorated. Many spores have a distinct print on their wall. But there are some rare cases where the colour of the spore is due to its cytoplasm. An example where the cytoplasm is responsible for spore print is Clavaria helicoids. Basidial And Basidiospore Development Ultrastructure And CytochemistryAlthough the basidium has been identified as a separate form of the reproductive cell since the early nineteenth century, there has been little attention to cytoplasmic processes in its development until recently. Corner's (1948) extensive light microscopic account of non-nuclear features of basidial growth offered a key incentive for future research and a theoretical framework. The study of the basidium's ultrastructure began in the 1960s. 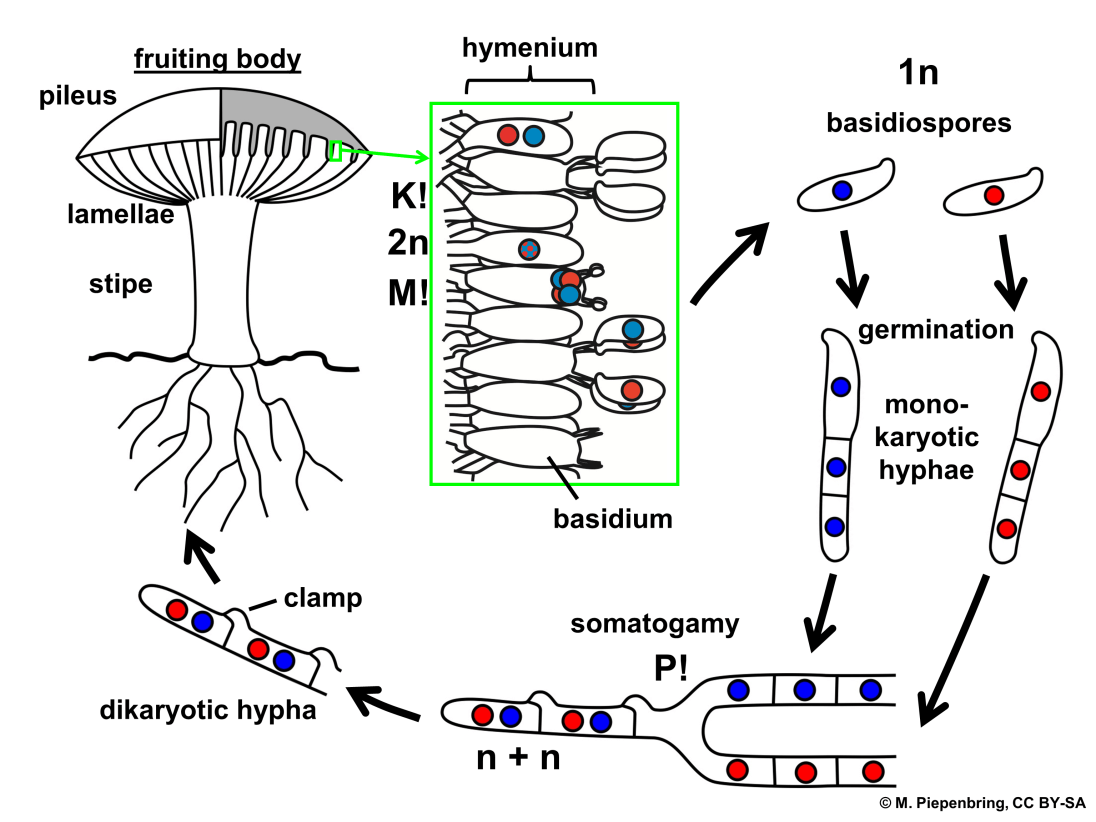
The majority of this research has focused on the holobasidium. However, only the Agaricales and Aphyllophorales have been thoroughly investigated. Even in Coprinus, possibly the best-researched genus, the cytoplasmic processes in holobasidial development are not entirely understood. Although there has been a significant rise in articles on phragmobasidial ultrastructure in recent years, a thorough explanation of the cytoplasmic processes in phragmobasidial formation remains absent. Basidiospore Discharge MechanismThe basidiospores in most Basidiomycota are ballistospores, which are aggressively released. The propulsive force is generated by a rapid shift in the spore's centre of gravity. The main factors responsible for forcible discharge are Buller's drop, attachment of the spore to sterigmata and the presence of a hygroscopic region on the surface of the basidiospores. The Buller's drop is a fluid droplet accumulated on the spore's hilar appendage or proximal region. The spore is attached to the tip of the sterigma through an offset attachment. Basidiospore discharge can occur only once enough water vapour has formed on the spore. When a basidiospore matures, sugars in the cell wall begin to function as condensation loci for water vapour in the air. Two distinct condensation areas are necessary. Buller's drop gathers as a huge, nearly spherical water droplet at the pointy tip of the spore (the hilum) closest to the supporting basidium. 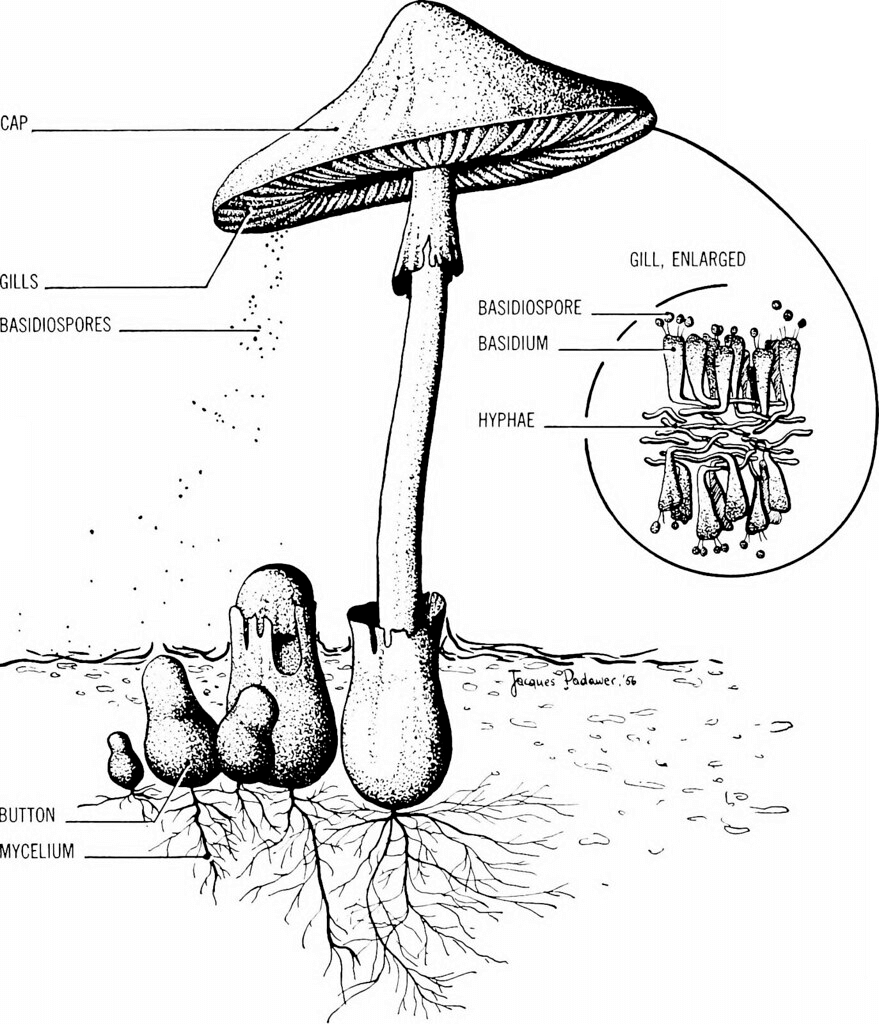
Condensation happens as a thin coating on the adaxial face of the spore at the same moment. When these two bodies of water collide, the release of surface tension and the abrupt change in the centre of mass cause the basidiospore to discharge. The spore's initial acceleration is expected to reach over 10,000 g. Discharge Of SporesSome basidiomycetes cannot eject their basidiospores but continue to generate them. Spore dispersion happens via distinct discharge mechanisms in each of these categories. As an example:
In these circumstances, the basidiospore usually lacks a hilar appendage, and there is no forced release. Each occurrence indicates a unique evolutionary loss of the basidiomycete forceful discharge mechanism.
Next TopicBlood Bank
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








