| |

Basal Ganglia PartsStriatumDeep inside the insular lobe of the forebrain's subcortical structures, there is a complex nucleus present which is known as the striatum. The striatum is divided into two different regions i.e., dorsal, and ventral regions. We will not go into greater detail about the ventral striatum because it is considered a limbic system component. On the other hand, the dorsal striatum is a part of the basal ganglia, and it is most often known as striatum. The caudate nucleus and putamen comprise the two components of the dorsal striatum, sometimes known as the striatum. The internal capsule, whose myelinated fibres extend into the striatum and give it a distinctive striped look, divides the components of the striatum. The striatum is a component of the corpus striatum, which also includes the globus pallidus. 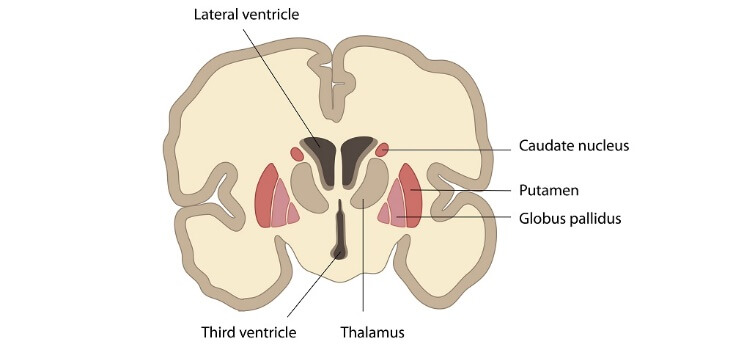
80-95% of the striatum's substance comprises projection neurons, which are small to medium-sized spiny neurons. The moniker "projection neurons" refers to the many spines that cover them. They operate as inhibitory neurons and release GABA into the nervous system. These neurons' axons are connected to the globus pallidus and substantia nigra via the route of direct and indirect basal ganglia. Interneurons in the striatum can be divided into four classes based on their absence of spines: Large a Spiny Cholinergic NeuronGABAergic neurons with the following characteristics: Parvalbumin-containing neurons; Somatostatin/nitric oxide synthase-containing neurons; and Calretinin-containing neurons. These neurons control the activity of the thalamus, SNc, and cerebral cortex. Caudate NucleusThe bottom of the caudate nucleus covers the inferior part of the lateral ventricle. It curves over the thalamus' ventral surface, penetrates the temporal lobe, and joins the amygdala at its conclusion. The putamen and the caudate nucleus are contiguous at the rostral end, and the nucleus accumbent surrounds it inferiorly. The range of capabilities discussed in the striatum includes the caudate nucleus's functions. Specifically, the thalamus gain all the information from the caudate nucleus about the necessary or important fine tunings of the motor response to those stimuli after the caudate nucleus integrates sensory information about the spatial position of your body. It also affects the posture of the body and the limbs, as well as the quickness and precision of directed motions. The caudate nucleus is used in different processes in addition to motor control, including memory, learning, language processing, emotions, goal pursuit, etc. PutamenLentiform nucleus (Nucleus lentiform) The forebrain's base is in a round structure which is referred as putamen. On the axial portion of the brain, it is the most lateral basal ganglia. The putamen surrounds the globus pallidus like a shell and extends rostrally and caudally. It is located at the back side of the globus pallidus and medially to the external capsule. The internal capsule divides it from the caudate nucleus, which encircles it. The medial medullary lamina is a thin band of whitish matter, which lies between the putamen and the globus pallidus. The main role of the putamen is to co-regulate with the motor functions and influence different types of learning, and it employs dopamine to perform its necessary functions. Nucleus accumbens and olfactory tubercleThere are two structures at the base of the forebrain: the nucleus accumbency and the olfactory tubercle. They make up input nuclei for the ventral tegmental region and are parts of the ventral striatum (VTA). Nucleus accumbens is located where the head of the caudate nucleus and the putamen joins in the rostral forebrain. But between the optic chiasm and the olfactory tract, ventral to the nucleus accumbens, is where the olfactory tubercle is situated. These structures, collectively known as the "limbic-motor interface," are crucial components of the "reward circuit" but are not involved in regulating movement. The brain's ventral tegmental region (VTA) contains dopamine neurons that become active when we engage in pleasurable behaviours (such as eating, using drugs, or having sex). Dopamine levels increases when these neurons are stimulated because they project to the nucleus accumbens and the olfactory tubercle. Globus pallidusThe putamen-medial paired subcortical structure known as the globus pallidus is made up of inhibitory GABAergic projection neurons that fire often and erratically on their own. The medial (internal) medullary lamina, a strip of white matter positioned vertically, separates it into external (GPe) and inner (GPi) segments. The internal capsule is in touch with the superior and medial sides of the globus pallidus. The globus pallidus and caudate nucleus are divided by the tablet. The substantia innominata and the hypothalamus both are joined to the globus pallidus in the anterior brain. It is situated close to the optic tract more caudally. The putamen and globus pallidus is also referred to as the lenticular nucleus because of their proximity and similar bean-like forms. Both the GPe and GPi are essential to modulate the motor programme, specifically in the direct and indirect routes. They both receive striatopallidal fibres, also referred to as Wilson's pencils, from the striatum, which provides inhibitory GABAergic input. The internal region of the globus pallidus receives fibres from the striatum as part of the motor loop's indirect pathway. In the meantime, the direct motor loop pathway includes threads that run from the striatum to the exterior of the globus pallidus. The pallidothalamic tracts are the globus pallidum output fibres. They separate into the thalamic fasciculus, lenticular fascicles, and ansa lenticularis. They make up the Forel's field as a whole. The globus pallidus and thalamic nuclei are connected by these connections. Subthalamic nucleusWithin the subthalamus are tiny biconvex paired structures called subthalamic nuclei (STN), also called Luys' bodies. The basal ganglia do not anatomically include the subthalamic nucleus. The subthalamus is a functional component of the basal ganglia due to its helpful link. The subthalamic nucleus is ventral to the thalamus and ventrolateral to the red heart at the intersection of the diencephalon and midbrain. It is covered by the medially in the internal capsule and anteriorly by the substantia nigra. The pallidothalamic fibres and the Forel's fields twine around the STN's ventral and medial boundaries before arching back over its dorsomedial surface, as the thalamic fasciculus is very strongly associated with it. Thus, these fibres isolate the zone incerta from the thalamus above and the subthalamic nucleus below. Neurons with excitatory glutamatergic projections make up the subthalamic nuclei. It gets somatotopically structured excitatory impulses from the frontal brain. This leads to the division of the subthalamic heart into three sections:
Substantia NigraSubstantia nigra, which is a small motor nucleus is in the midbrain's anterior region, between the cerebral peduncle and tegmentum. Despite being in the midbrain, it is a functional basal ganglia component. The pars compacta (SNc) and the pars reticulata are the two components that make up the substantia nigra, and their connections and functions are highly distinct (SNr). From both sides of the midbrain, it separates the cerebral peduncles from the tegmentum. The red and subthalamic nuclei border it dorsomedial, and the medial lemniscus and geniculate bodies connect it laterally. The substantia nigra's dorsal section is made up of the pars compacta. The substantia nigra's unique dark hue results from the large number of densely packed melanin-filled neurons that make up this structure. In front of the pars compacta is the pars reticulata. Although it is bigger than the pars compacta, it has fewer cells. The ventral tegmental area is a region that lies medially to the substantia nigra. The pars compacta may be considered an extension of this section because it is a small collection of dispersed cells with identical functions. Through particular D1 and D2 neurons in the nigrostriatal pathways, the pars compacta primarily function as an output to the basal ganglia circuit, feeding the striatum with dopamine. However, the pars reticulata mainly serves as an input, sending information to the thalamus from the basal ganglia. Parkinson's disease and several other parkinsonism disorders are thought to occur due to the death of dopamine neurons in the SNc.
Next TopicBlood Cells
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








