| |

Bacterial ShapesDespite the fact that bacterial cells are significantly smaller and have a simpler structure than eukaryotic cells, bacteria are a varied collection of creatures.They vary in size, shape, environment, and metabolism. Many of our current understandings of bacteria derive from studies of disease-causing bacteria, which are easier to isolate in pure culture and analyze than many free-living bacterium species. It's worth noting that many free-living bacteria aren't the same as bacteria that have evolved to live as parasites or symbionts on animals. As a result, there are no absolute principles for bacterial composition or structure, and each general statement is subject to many exceptions. 
Bacteria may have one of three basic shapes: spherical (coccus), rodlike (bacillus), or curved (bacillus) (vibrio, spirillum, or spirochete). The actual forms of bacteria show a lot of diversity, and cells may be stretched or compressed in one dimension. Bacteria that do not split from one another during cell division produce distinct clusters that may be used to identify them. Some cocci, such as Streptococcus pneumoniae, pneumococcus that causes bacterial lobar pneumonia, and Neisseria gonorrhoeae, a gonococcus that causes the sexually transmitted illness gonorrhea, are mainly found in pairs. The staphylococci form random clumps, while the streptococci resemble a long thread of beads (the term "staphylococci" comes from the Greek word staphyle, which means "cluster of grapes"). 
Furthermore, certain coccal bacteria may be found in a square or cubical packages. The rod-shaped bacilli are normally found individually, but other strains, such as rods of the corynebacteria, which are common oral residents and commonly adhere to one another at odd angles, create lengthy chains. Other bacilli have pointy endpoints, while others have squared ends, and some rods have a comma shape. Vibrio cholerae, which causes cholera, is a type of twisted rod known as vibrios. The spirilla, which are bent and rebent, and the spirochetes, which form a helix akin to a corkscrew, with the cell body wrapped around a central fiber termed the axial filament, are two more types of bacteria. Questions regarding how a cell creates a rod form have dominated the study of bacterial morphology, which is, of course, only one example of the more general subject of how a cell constructs any shape. It is hoped that by answering this (deceptively) easy question, we will get information that will lead us to a universal shape control mechanism. This is logical given that we are more experienced and comfortable addressing how-type queries. And as other papers in this issue demonstrate, this technique has yielded interesting new knowledge. 
Every Shape Has a Distinct ValueThe first point to establish is that a bacterium's form has biological significance. One argument in support of this claim is that, despite the huge diversity of forms available to bacteria, each specie often shows just a small subset of them, implying that, given a universe of shapes to select from, individual bacteria choose only those that are adaptable. Another hint is that certain bacteria may change their morphology in response to environmental signals or during pathogenesis, implying that form is essential enough to warrant control. Two evolutionary theories further support the value of the bacterial form. The earliest shape has a vector across evolutionary time, with rodlike animals emerging first and coccoid species appearing as derivatives at the ends of evolutionary lines. The progression of a character suggests that selection factors are at work. Second, prokaryotes from various lineages may morphologically converge, implying that having a similar form may provide benefits in some situations. The Archaea, for example, have a non-peptidoglycan-based cell wall yet display a spectrum of morphological forms that are comparable to Bacteria. The most straightforward conclusion is that morphological adaptation serves a vital biological purpose. 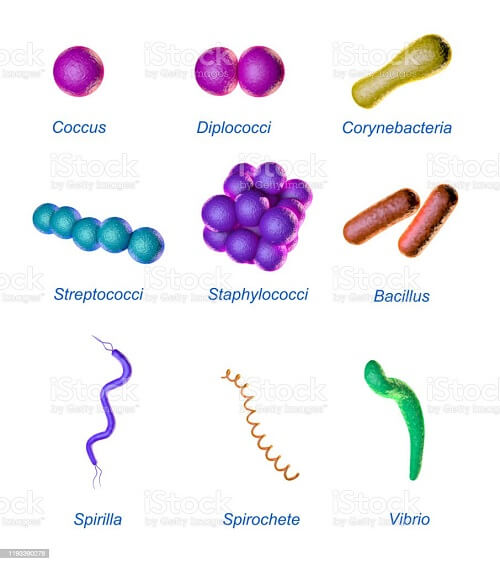
So, how does morphology play a role in natural selection? Simply said, bacteria of various shapes exhibit distinct physical characteristics to the outside world and these characteristics aid cells in coping with and adapting to environmental situations. Even a 0.01 percent increase in E. coli growth rate confers a fitness advantage of 10% over its unaffected rivals, indicating that improvements do not have to be spectacular to be helpful. Shape contributes a measure of survival value in the face of three "Primary" selective pressures: nutrient acquisition predation and cell division. The shape of the cell also contributes to the optimization of 5 secondary mechanisms: passive dispersal, internal or external differentiation, active motility, and attachments to surfaces. Since the cells must grow and increase while avoiding being destroyed, the first three are primary in that they reflect basic circumstances that decide whether cells survive or die. The remaining five are secondary in that they constitute a group of morphologically related methods that bacteria use to combat the primary forces. Read on to explore the three of them and to know how selective pressures affect cell shape. Nutrient AbsorptionThe conventional response to the issue of why prokaryotic cells are so tiny is that they need a significant surface-to-volume ratio to maintain their internal biochemistry. Koch calculated how big a cell might get if it only split once a day, like the massive symbiotic bacteria Epulopisciumfishelson, and relied exclusively on diffusion in a nutrient-rich environment. His response was that a bacteria might have a diameter of almost 800 meters! This means that the size restrictions of more typical prokaryotes are caused by competition for resources, which is won mostly by smaller, faster-growing cells. The lesson is that, although diffusion-limited nutrition supply may influence cell size, it does not explain why bacteria are usually tiny on their own. On the other hand, diffusion considerations explain how bacteria might improve their nutrient collecting efficiency by changing their gross shape. One of the researchers recently published a paper that backs up the concept that this modest change in cell shape is a physiologically valuable response. They proved the presence of stalk-mediated phosphate absorption and mathematically showed that extruding a long thin stalk instead of filamenting allows cells to absorb more phosphate. This is a substantial experimental proof that cell form influences nutrient uptake in and of itself, and it suggests that different nutritional circumstances may prefer one bacterial shape over another. MotilityAll kinds of movement, in theory, exert high physical and energy demands on cell structure. The energy needed for chemotaxis may be changed by a factor of 105 with a change in cell diameter of just 0.2 m! By far, the well-studied morphological link is that between bacterial shape and motility. Highly motile bacteria are forced into a small range of ideal sizes and rod forms by energy consumption, Brownian forces, and the need to follow chemical gradients. The behavior of filamentous E. coli cells, although motile and chemotactic, travel slowly and cannot tumble to change direction, supports these theoretical concerns. Cells that move as a group rather than as individuals are subject to different morphological restrictions. Since the group aligns itself via significant side-by-side cell-to-cell interactions, such "swarm cells" are longer than single cells. Certain Proteus mirabilis mutants become non-motile due to their extremely curved swarmer cells, which are unable to align correctly due to the shapeshift. 
In a nutshell, movement puts a lot of strain on cell structure. If we think logically, then it should be rod-shaped bacteria that are fast with a certain length to ratio. And it should be chemotactic cells that adopt a shape that matches their habitats. Also, it should be spiral or curved-shaped cells that graze near surfaces or negotiate viscous environments. PredationOne of the least known evolutionary stresses that bacteria undergo is Predation by protozoa, commonly known as protistan grazing or bacterivory. Predation is one of the most evident selection pressures impacting bigger (i.e., non-microbial) creatures. Therefore this lack of awareness is unexpected. Several reviews have attempted to address this gap. An example of top-down selection is Protistan grazing, where external organisms give evolutionary pressure. On the other hand, bottom-up selection includes food availability and cell division because they impact the organism's cells through basic reproduction needs. Bacteria evolve escape mechanisms in response to Predation, resulting in a well-known arms race between predator and prey that leads to bacterial diversity. 
Cell shape influences three fundamental defensive strategies:
All of these are influenced by one or more features of bacterial morphology, either directly or indirectly. Today, a lot of information is available, along with the experimental evidence that the selection of cells that can change their form or size is done on grazing when there was earlier just conjecture. Protistan feeding causes bacteria to become tiny or extremely big, move quicker, filament, generate prosthesis, proliferate as microcolonies, become longer or curved, filamentous or chained, or increase their diameter. The cyanobacterium Arthrospira, which develops as helical trichomes, is one of the most fascinating morphological changes. A ciliate feeding on this creature spins on its long axis to consume up to six entire coils. Arthrospira, on the other hand, may modify its helical handedness from right to left or vary the pitch of its spirals to minimize Predation. In brief, bacteria have developed morphological barriers in their fight against being eaten, which may have resulted in the variety of morphologies we see today. Summary and ComplexitiesEnvironmental factors function in tandem, eliciting a wide range of reactions. This makes biological prediction a hazardous endeavor at best, and it's more dangerous when it comes to cell shape since many selection pressures may drive morphological change in unanticipated ways. During a nutritional shortage, a rod-shaped cell lacking prosthecae (dark blue) becomes tiny and coccoid to preserve energy and avoid predator capture. The stalk of a Caulobacter-like cell (light blue) aids in nutrient harvesting during times of shortage and prevents swallowing when predators are plentiful. The cells in this example change form in one or two generations, while other species or situations may need acclimation over time. It's simple to understand how adding more selection pressures and taking into account more morphological responses might result in a broad range of shape optima for dealing with various situations. We know very few morphological laws, notwithstanding some fundamental patterns (for example, motile cells are generally rods). Consider the bacteria Pelagibacterubique, which accounts for almost a quarter of all ocean microorganisms and is arguably the most successful and widespread single prokaryote. We have no idea why it is a little curved rod instead of a small straight rod, even for such a simple bacteria in such a straightforward environment, and beyond that, everything else is much more unknown. In summary, with our current level of knowledge, we cannot anticipate an organism's form from its surroundings, nor can we accurately deduce the properties of its environment from its shape. In conclusion, appearance isn't everything. The idea is that morphology is an important selected feature in many situations and can be studied experimentally in the same way as any other. We may soon be able to alter bacterial morphology with enough certainty to investigate how morphological changes impact survival in diverse settings as we learn more about the systems that govern cell shape. As more evidence demonstrating the value of cell shape accumulates, we may expect that researchers will be more encouraged to pursue these sorts of issues directly.
Next TopicPhotosynthetic bacteria
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








