| |

Sexual Reproduction in Flowering PlantsHuman beings have enjoyed an intimate connection with flowers since the beginning of time. Flowers are aesthetic or ornamental, religious, social, and symbolic values. They have been used for centuries to convey basic human emotions like the love of a loved one, joy and sorrow, and so on. 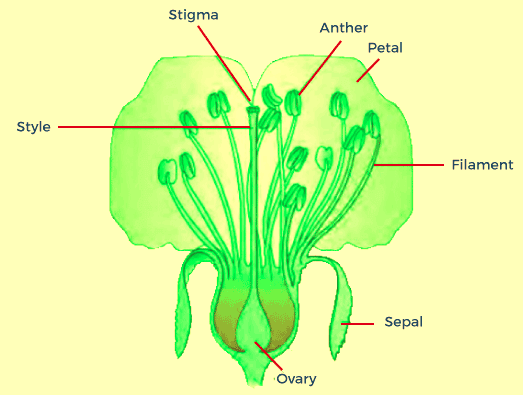
PRE-FERTILISATION - STRUCTURES AND EVENTSPre-fertilization simply means the events that take place before fertilization. Prior to the moment, the actual flower appears on an individual plant, the plant's choice to flower has already been taken. A variety of hormone and structure changes occur that result in the differentiation and subsequent growth of the primordium of flowers. Inflorescences grow, which produce the floral buds and later the flowers. The reproductive systems of males and females in flowers, including the androecium and the gynoecium, both differentiate and grow. The male part, i.e., androecium, consists of stamens, while the gynoecium symbolizes that female organ for reproduction which consists of stigma, style, and ovary. Stamen Microsporangium, and Pollen Grain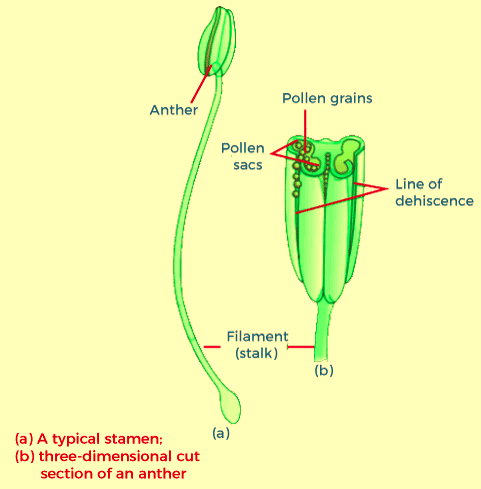
These diagrams depict that the two main components of the common stamen- the lengthy and slender stalk is called the filament, and the bilobed structure typically formed at the terminal is known as the anther. The proximal part of the filament connects to the thalamus or the petals in the bloom. The length and number of stamens differ in the flowers of various species. If you took ten stamens from flowers (each of various species) and then arranged the stamens on slides, you'll be amazed after seeing the considerable variation in size of pollen grains that are seen in the natural world. A careful examination of every stamen under an optical microscope and then creating precise diagrams will help understand the different shapes and anthers' attachment in various flowers. Angiosperm anthers are bilobed, with each lobe having two thecae, i.e., these can be described as dithecous. Often, a longitudinal groove runs across the length of the theca, which separates the theca. The bilobed structure in an anther can be distinctive in the transverse section that makes up the anther. The anther is a quadrilateral (tetragonal) structure made up of 4 microsporangia situated in the corners and two per lobe. The microsporangia grow further and eventually become pollen sacs. These sacs further extend along the length of the anther, and later they get packed with pollen grains. Microsporangium's Structure: The typical microsporangium appears close to circular in shape in a transverse section. It is usually covered by four wall layers: the epidermis, the endothecium, the middle layers, and the tapetum. The three outer wall layers play the role of protecting and aiding in the dehiscence of the anther to let the pollen out. The wall's innermost layer is called the tapetum, and it is the source of nutrition for emerging pollen grains. The tapetum cells have abundant cytoplasm and usually contain multiple nuclei. If the anther is still young, the anther is composed of compactly laid out homogenous cells known as the sporogenous tissue located in the middle of each microsporangium. Microsporogenesis: When the anther grows, the cells in this sporogenous organ undergo meiotic divisions that create microspore tetrads. Each cell in the sporogenous tissue can produce microspores in a Tetrad. Each of them is a potential microspore or pollen mother cell. The process of forming microspores in a pollen mother cell (PMC) by meiosis is known as microsporogenesis. Microspores, when they develop, are placed in a group of four cells known as the microspore tetrad. Within each microsporangium, many pollen grains or microspores produce and are released during the breaking of the anthers. When the anthers grow and begin to dehydrate, the microspores separate and transform to form pollen grains. Pollen grains: Pollen grains symbolize the male gametophytes. If you touch the open anthers of Hibiscus or another flower, you will see an accumulation of yellowish, powdery grains on your fingertips. Sprinkle the grains onto the drop of water onto a glass slide to examine them under the microscope. You'll be amazed by the range of different architectural forms - sizes, forms, colours, and patterns visible on pollen grains of other species. The pollen grains are usually round, measuring between 25 and 50 millimetres across. It is distinguished by its two-layer wall; the tough outer layer is known as "exine." The exine is composed of sporopollenin, which is the most durable organic material recognized, and it can withstand extreme temperatures and strong acids and alkali. No enzyme can degrade sporopollenin, or we can say it is yet to be identified. Exine of the pollen grain has noticeable openings, also known as germ pores, in which sporopollenin is not present. The pollen grains are preserved as fossils due to the presence of the sporopollenin. 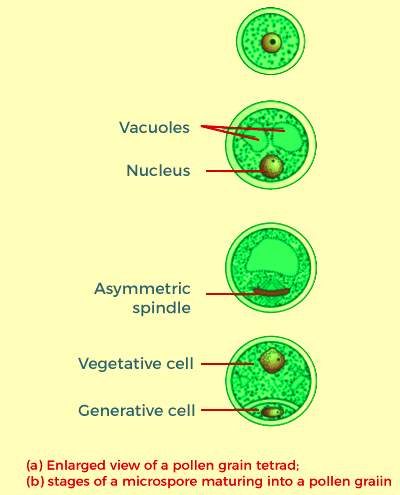
Exine displays an intriguing assortment of designs and patterns. The inside of the pollen grain is referred to as the intine. This layer is an extremely thin, continuous layer of pectin and cellulose. The plasma membrane encloses the cytoplasm of the pollen grain. If the pollen grain is mature, it has two cells: the vegetative and generative cells. The vegetative cell is more extensive and has massive food storage and a nucleus with an irregular shape. The generative cell is tiny and floats in the cytoplasm of the vegetative cell. It has a spindle shape, with dense cytoplasm and nucleus. More than 60 percent of angiosperms shed the pollen grains in a two-celled stage. In the other species, the cells that generate the pollen divide mitotically to produce two male gametes before the pollen grains are removed (three-celled the stage). Pollen grains from many species can trigger severe allergic reactions and bronchial problems in certain patients, frequently creating chronic respiratory problems such as asthma, bronchitis, etc. It is possible to mention that carrot grass, or Parthenium introduced in India as a contaminant from imported wheat, has become widespread in its distribution and can cause pollen allergies. Pollen grains are rich in nutrients. In recent times it's been a trend to use pollen tablets for food supplements. In Western countries, there are a lot of pollen products in syrups and tablets readily available on the market. The consumption of pollen has been proven to boost the athletic performance of athletes and racehorses. After shedding, the pollen grains need to settle upon the stigma before losing their viability to fertilize successfully. What is the length of time you believe the pollen grains will last? The time frame during which pollen grains can remain viable is a bit dependent and, to a certain degree, is contingent on the current temperatures and humidity. The pollen grains of certain cereals, such as wheat and rice, die within 30 minutes after their release. In other species of Rosaceae, Leguminosae, Rosaceae, and Solanaceae, they remain viable for months. There is a possibility of storing sperms and semen from numerous animals, including humans, to induce artificial reproduction. There is a way to keep pollen grains of a wide variety of species over time with Liquid Nitro (-196o). These stored pollen grains can create pollen banks like seed banks in breeding crops. The Pistil, Megasporangium (ovule), and Embryo sacThe gynoecium is the female reproductive component in the bloom. The gynoecium could be composed of one pistil (monocarpellary) or multiple pistils (multicarpellary). If more than one pistil, they could be joined (syncarpous) or completely free (apocarpous). Each pistil comprises three parts: the stigma, style, and ovary. The stigma acts as an entry point that pollen grains can land on. Style is the long and slender part that lies beneath the stigma. The part that bulges at the base of the pistil constitutes the ovary. Inside the ovary, there is a cavity of the ovarian (locule). The placenta is in the ovarian cavity. The megasporangia that arise from the placenta are often referred to as Ovules. Many ovules within an ovary could range from one (wheat paddy, mango, or wheat) to several (papaya watermelon, papaya orchids). The Megasporangium (Ovule): Let us get acquainted with the basic structure of the angiosperm's ovule. The ovule can be described as a small structure attached to the placenta with the help of a stalk, also known as a Funicle. Its body joins with the funicle in an area known as the hilum. The hilum is the intersection between the ovule and funicle. Each ovule contains two or more protective envelopes known as integuments. They surround the nucellus, except when a tiny opening known as the micropyle is arranged. At the opposite end of the micropylar, there is an area called the chalaza, which is the base of the ovule. There is an array of cells in the integuments known as the nucellus. The nucellus cells have an abundance of reserve food sources. The nucellus contains its egg sac or female gametophyte. An ovule typically has an embryo sac made by the megaspore. 
Megasporogenesis: A process that involves the formation of megaspores out of megaspore mother cells is known as megasporogenesis. Ovules are the most common way to differentiate one megaspore mother cell (MMC) within the micropylar part within the nucellus. It is a massive cell with dense cytoplasm and an imposing nucleus. The large size and prominence of the nucleus characterize MMCs. MMC is undergoing meiotic division, and meiosis causes the creation of 4 megaspores. Gametophyte female: In the most flowering plant, one megaspore is functional, while the remaining three are degenerate. The only functional megaspore develops into the female gametophyte (embryo sac). This method of forming an embryo sac by a single megaspore is known as monosporic development. Let's examine the development of an embryo sac in more detail. Megaspores that function as nuclei split mitotically into two nuclei moved to opposite poles, creating an embryo sac that is a two nucleated embryo sac. Two additional mitotic nuclear divisions cause the creation of the four-nucleate and, later, in the eight-nucleate stage of the embryo sac. It is essential to know that these mitotic divisions are free nuclear. That is, they are not followed immediately by the formation of cell walls. After the 8-nucleate stage, cells begin to form, and the process begins to form an average female gametophyte or embryo sac. Examine the arrangement of cells within the sac of embryos. The six nuclei have been surrounded by cell walls and are organized into cells. The remaining two nuclei, referred to as Polar nuclei, are found beneath the egg apparatus within the big center cell. There is a distinct pattern of cells in the sac of embryos. Three cells are placed at the micropylar tip and make up an egg's apparatus. The egg apparatus comprises two synergids as well as an egg cell. Synergids are characterized by cells that thicken at the tip of the micropylar, known as the filiform apparatus. These have a crucial role to play in steering the pollen tubes to the synergid. Three cells reside at the chalazal edge and are known as antipodals. The central cell is the largest, like the one mentioned earlier contains two polar nuclei. So, a typical angiosperm embryo sac at the time of maturation, although eight nucleates, is seven-celled. PollinationIn the earlier sections, we've been taught that female and male gametes of flowering plants are non-motile, so they must be joined for fertilization to take place. Pollination is the method used that achieves this aim. Pollen grain transfer (shed out of the anther) to the pistil's stigma is known as pollination. The flowering plants have evolved numerous methods to facilitate pollination. DOUBLE FERTILISATIONWhen it enters one synergid, the pollen tube releases two male gametes released into the cell cytoplasm of the synergid. The male one move toward the egg cell and then fuses with the nucleus, thus concluding the syngamy. This causes the formation of diploid cells, the zygote. The male gamete then moves toward the two nuclei in the central cell and then fuses with them to form a primary endosperm nucleus that is triploid (PEN). Because this involves the combination of three haploid nuclei, it is called triple fusion. Since there are two kinds of syngamy and triple fusion, which occur in embryo sacs, the process is known as double fertilisation. This uncommon event occurs only in flowering plants. The central cell that is formed after triple fusion transforms into its primary endosperm cells (PEC) and then develops into the endosperm. The zygote grows into an embryo. POST-FERTILISATION: STRUCTURES and EVENTSFollowing double fertilisation, occurrences of the development of embryos and endosperm development of ovule (s) into the seed(s) and the ovary to fruit are collectively referred to as post-fertilisation processes. EndospermEndosperm development precedes embryo development. The primary endosperm cell divides in a series of cycles and forms a triploid tissue. The cells in this tissue are stuffed with foodstuffs that are stored in reserve and serve to provide nourishment for the embryo in development. In the typical type of development of endosperm development, the PEN undergoes a series of nuclear divisions, leading to free nuclei. This stage of development is known as the free-nuclear endosperm. Then, cell wall formation occurs, and the endosperm becomes cellular. The number of nuclei that are free created before cellularisation is different. The coconut water we drink from the tender coconut, which you're familiar with, is nothing more than free-nuclear endocrine (made by many cells) and the white kernel constitutes the cell-based endosperm. The developing embryo's endosperm could be either consumed completely (e.g., beans, groundnut, pea) before the maturation of seeds or may remain in persistent seeds (e.g., coconut and castor) and then be utilized in the process of seed germination. EmbryoThe embryo develops at the micropylar tip in the sac of the embryo, in which the zygote is located. The majority of zygotes split after an amount of endosperm has been produced. This is an adaptation that helps ensure the nutrition of the embryo in its development. While the seeds differ in size initially, the stages of the embryo's development (embryogeny) are the same for dicotyledons and monocotyledons. The zygote is the source of the proembryo, which then leads to the heart-shaped, rounded, and mature embryo. A typical dicotyledonous sperm consists of an embryonal axis and two cotyledons. The embryonal axis that is above those of cotyledons is the epicotyl. It ends with the stem or plumule end. The cylindrical part below that level is hypocotyl, which ends at the lower part of the radicle, or root tip. It is covered by the root cap. Monocotyledon embryos only have one cotyledon. Within the family of grasses, the cotyledon is known as the scutellum and is located on the opposite of the sides (lateral), the embryonal axis. Its lower end is where the embryonal axis comprises the root cap and radical wrapped in a sheath that is not differentiated, known as coleorhiza. The embryonal axis, which is higher than the attachment point of the scutellum, is known as the epicotyl. Epicotyl shoots at the apex, and some leaf primordia enclosed within a hollow foliar form the coleoptile. SeedIn angiosperms, it is the ultimate result of the sexual cycle, and it is usually described as fertilised ovule. The seeds are produced inside the fruits. A seed usually consists of the seed coat(s), cotyledon(s), and an embryo axis. The cotyledons in the embryo are simple structures that are generally massive and thick because of the storage of reserves for food (as for legumes). Mature seeds can be ex-albuminous or non-albuminous. Non-albuminous seeds do not have any residual endosperm since it is consumed during the embryo's development (e.g., pea groundnut). Albuminous seeds contain a small amount of the endosperm since it is not entirely consumed during the development of sources (e.g., wheat, maize, or castor). Sometimes, nucellus remnants remain in certain seeds like black pepper or beet. This persistent, residual nucellus can be found in the perisperm. The ovules' integuments become strong seed coats. The micropyle is a tiny pore within the coat of seeds. This facilitates the entry of water and oxygen into the seed during the germination process. When the seed matures, its water content decreases, and seeds turn dry (10-15 percent moisture in mass). The metabolic activity in general of the embryo decreases. The embryo could go into a state of no activity called dormancy. If favorable conditions are present (adequate humidity, oxygen levels, and adequate temperature), the embryos begin to grow. When ovules develop and become seeds, the embryo transforms into a fruit, i.e., the process of turning ovules into seeds and the ovary into fruit is happening simultaneously. The ovary's wall transforms into the fruit's wall called the pericarp. The fruit may be soft, like mango, guava, orange, etc. Or they could be dry, as in mustard, groundnut, and others. Many fruits have developed mechanisms to disperse seeds. Remember the different types of fruits and their mechanisms for dispersal you studied in a previous class. When the fruit emerges from the ovary in all plants, additional flowering parts begin to degenerate and disappear; however, in some species, such as apples or strawberries, cashew, and so on. The thalamus helps in the formation of fruit. These fruits are referred to as false fruits. They are mainly produced out of the ovary and are referred to as real fruits. In the majority of species, fruits are the result of fertilisation. However, there are some species where fruits grow without fertilisation. They are known as parthenocarpic fruit. Banana is an instance. Parthenocarpy can be stimulated through the use of growth hormones. Such fruits are seedless. Seeds have numerous advantages over angiosperms. First, reproductive processes like fertilisation and pollination are not dependent on the presence of water. Seed production is more stable. Secondly, seeds possess better strategies to adapt to new habitats and help the species. To expand into new areas. Because they have enough food sources, the seedlings get enough nutrition until they can self-photosynthesis. The hard seed coat protects the embryo. As products from sexual reproduction, seeds create new genetic combinations that lead to different varieties. Seed is the base of our farming. Dormancy and dehydration in mature seeds are essential in storing seeds so that they are used for food throughout the year and increase crop next year. This time period is also very variable. For a handful of species, seeds die within several months. Seeds from a wide range of species last for several years. Certain seeds can live over hundreds of years. There is considerable evidence of extremely old but viable seeds. The oldest one is Lupine (Lupinus arcticus), discovered in Arctic Tundra. The seed germinated and then blossomed after an estimated 10000 years of dormancy. The most recent evidence of 2000 years old and viable seeds is part of the date palm (Phoenix Dactylifera), discovered during the archeological excavations at King Herod's palace close to the Dead Sea. APOMIXIS AND POLYEMBRYONYWhile seeds generally are products of fertilization, some flowers, like certain kinds that belong to Asteraceae and grasses, have developed a unique method of producing fertilized seeds, referred to as the apomixis. Apomixis is a kind of sexual reproduction that replicates sexual reproduction. There are many ways to the development of apomictic seeds. The diploid egg develops without reducing division in certain species and grows into various varieties. Nucellar cells around the embryo sac begin to divide in the embryo sac and then develop into embryos. In these species, each ovule is home to several embryos. Without fertilization, a single embryo in one seed is known as the polyembryony egg. Most often, in several Citrus and Mango varieties, a few of the nucellar cells that surround the embryo sac begin to divide and then protrude inside the embryo sac and grow into embryos. In these species, each ovule is home to numerous embryos. The presence of more than one embryo within a seed is known as polyembryony. Hybrids of various vegetable and food crop varieties are widely cultivated, and the cultivation of hybrids has dramatically improved productivity. One of the issues with breeding hybrids is that hybrid seeds need to be produced annually. If the seeds from hybrids are planted, the progeny seeds can segregate and lose their hybrid traits. The production of hybrid seeds is expensive, and the costs of hybrid seeds are too costly for farmers. If the hybrids are bred into apomicts, there is no segregation of the characters within the progenitors of hybrids. Farmers can continue using hybrid seeds to grow new crops each year and are not required to purchase hybrid seeds each year. Due to the significance of apomixis in the hybrid seeds, research is being conducted in numerous labs around the globe to study the genetics behind apomixis and transmute apomictic genes to hybrid varieties.
Next TopicBryophyte
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








