| |

Plastid DefinitionA membrane-bound organelle present in the cells of plants, algae, and certain other eukaryotic creatures is called a plastid (derived from the Greek word plast�s meaning "formed, moulded"; the plural is "plastids"). They're thought of as intracellular endosymbiotic cyanobacteria. Examples include leucoplasts, which are non-pigmented plastids with the ability sometimes to differentiate; chloroplasts, which are used for photosynthesis; and chromoplasts, which are used for pigment production and storage. Around 1.5 billion years ago, an event that resulted in permanent endosymbiosis in the Archaeplastida clade of land plants, red algae, and green algae most likely involved a cyanobiont (symbiotic cyanobacteria) related to the genus Gloeomargarita. A later main endosymbiosis event occurred in photosynthetic Paulinella amoeboid between 90 and 140 million years ago. This plastid is a member of the Prochlorococcus and Synechococcus cyanobacterial genus "PS-clade" of plastids. In addition, some species sequester ingested plastids in a process known as kleptoplasty, and secondary and tertiary endosymbiosis have also been seen in a broad range of taxa. The term "plastids" and a precise definition were originally coined by A. F. W. Schimper. The kind of pigments in a plastid determines the colour of the cell, and they often include pigments needed in photosynthesis. They are also where significant chemical compounds that are used by the cells of autotrophic eukaryotes are produced and stored. They have a circular double-stranded DNA molecule, similar to prokaryotic organisms' circular chromosomes. The plastid is retained even in species when its photosynthetic abilities have been lost because of its crucial function in the synthesis of chemicals like isoprenoids. OriginThought to be endosymbiotic cyanobacteria are plasmids. Eukaryotes were able to perform oxygenic photosynthesis thanks to the main endosymbiotic event of the Archaeplastida, which is thought to have happened roughly 1.5 billion years ago. Since then, three evolutionary lineages of the Archaeplastida have appeared, each with a unique name for the plastids: chloroplasts in green algae or plants, rhodoplasts in red algae, and muroplasts in glaucophytes. The colouration and ultrastructure of the plastids are different. For instance, plants and green algae's chloroplasts no longer include any phycobilisomes, the light-harvesting structures present in cyanobacteria, red algae, and glaucophytes in their place. Instead, they now only possess stroma and grana thylakoids. Contrary to chloroplasts and rhodoplasts, the glaucocystophycean plastid is still encircled by the cyanobacterial cell wall. Two membranes enclose each of these main plastids. It is the only known primary endosymbiosis event of cyanobacteria outside of Archaeplastida. The plastid of photosynthetic Paulinella species, often known as the "cyanelle" or "chromatophore," experienced a considerably more recent endosymbiotic event around 90-140 million years ago. The plastid is a member of the "PS-clade" (of the cyanobacterial genera Prochlorococcus and Synechococcus), which is not the same as the sister clade that includes the plastids that are members of the Archaeplastida. Contrary to primary plastids, which come from a prokaryotic cyanobacteria's primary endosymbiosis, complex plastids come from secondary endosymbiosis, in which one eukaryotic organism engulfs another eukaryotic organism that has a primary plastid. The algal plastid is often encircled by more than two membranes when a eukaryote consumes a red or green alga and keeps it. These plastids' ability for metabolism and photosynthetic activity may be diminished in various circumstances. The majority of dinoflagellates (rhodoplasts), haptophytes, cryptomonads, and heterokonts are algae with complex plastids generated from the secondary endosymbiosis of a red alga. Euglenoids and chlorarachniophytes (chloroplasts) are among those that endosymbioses a green alga. Plasmodium spp., Toxoplasma gondii, and many other pathogens that cause human or animal diseases are all members of the Apicomplexa, a species of obligate parasitic protozoa that also contains a complex plastid (although some apicomplexans, like the cryptosporidiosis-causing Cryptosporidium parvum, have lost this organelle). Even though the 'apicoplast' can no longer perform photosynthesis, it is still a vital organelle and a promising target for the creation of antiparasitic drugs. Some sea slugs and dinoflagellates, particularly those of the genus Elysia, consume algae as food and retain the digested algal plastid in order to benefit from photosynthesis; eventually, the plastids themselves are digested. From the Greek word kleptes, meaning thief, this procedure is known as kleptoplasty. Types of PlastidsBased on their colour, shape, and ultrastructure, plastids may be classified into a number of kinds. Each plastid type's properties are closely tied to the functions that they play. The term "proplastids" refers to undifferentiated plastids, which are mostly found in meristematic and reproductive tissues. They may be distinguished from other plastid types by being tiny and possessing distinct ultrastructures. They can be distinguished as "leucoplasts" in white, "chloroplasts" in green, and "chromoplasts" in either yellow, orange, or red. "Etioplasts" refers to intermediate chloroplast forms, and "gerontoplasts" refers to senescent chloroplast forms. Leucoplasts are classified by their lack of colour. Still, they may also be further divided depending on their contents, such as "amyloplasts" that are starch-enriched, "proteinoplasts" that are protein-enriched, and "elaioplasts" that are lipid-enriched. 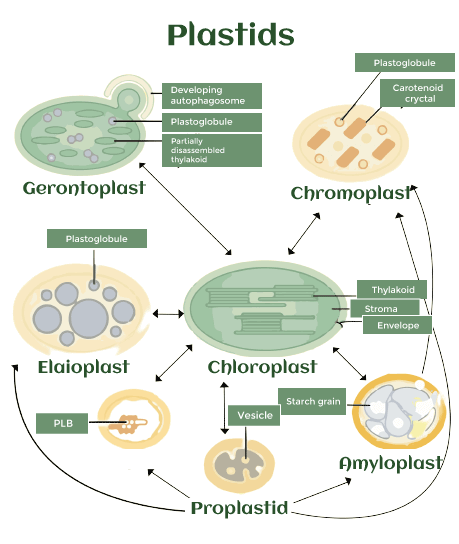
1. ProplastidsUndifferentiated plastids, known as proplastids, retain a basic plastid structure for the purpose of intergenerational organelle transfer. When compared to other types of plastids, they are colourless, very small, and lack any distinguishing morphological traits. They are mostly discovered in the meristematic and egg cells of plants, as well as sometimes during the generation of pollen in certain species like Pelagonium and barley. Additionally, it has been suggested that the nodule proplastids in root tissues are crucial to the biochemistry of nitrogen fixation in the legume family. 2. ChloroplastsAll organisms that engage in photosynthetic growth contain chloroplasts, one of the most extensively researched plastid types. Through the use of photosynthetic protein complexes, they can convert light energy into chemical energy. In chloroplasts, huge lipid surface layers known as grana are created by several stacks of thylakoids, which are single lipid layers that resemble discs and are used to attach photosynthetic protein complexes. Plastoglobules, which are distinct hydrophobic pocket structures formed on the borders of the disk-shaped thylakoids, contribute to increasing the interior area of the lipid bilayer. In order to produce apocarotenoids and accumulate non-endogenous carotenoids, plastoglobules are identified as sites of carotenoid breakdown. Chlorophyll, a key component of photosynthesis, gives chloroplasts their characteristic green colour. However, chloroplasts also contain large amounts of other hydrophobic terpenes that enable photosynthesis, including lutein, beta-carotene, violaxanthin, and neoxanthin. The carotenoid in chloroplasts not only convert UV-blue light into the electrical energy needed for photosynthesis, but they also significantly contribute to photoprotection by controlling the non-radiative dissipation of excess excitation energy. Particularly, hydroxylated carotenoids, also known as xanthophylls, aid in photoprotection by either directly quenching the triplets of chlorophyll (Chl) that are produced during photosynthesis or by scavenging the reactive oxygen species (ROS) that are produced. As a result, one of the key roles of chloroplasts is to maintain a balance between photosynthesis, photoprotection, and ROS scavenging. For temporal storage and to assist in responding to developmental and environmental signals, chloroplasts often generate starch granules, protein bodies, and lipid bodies. 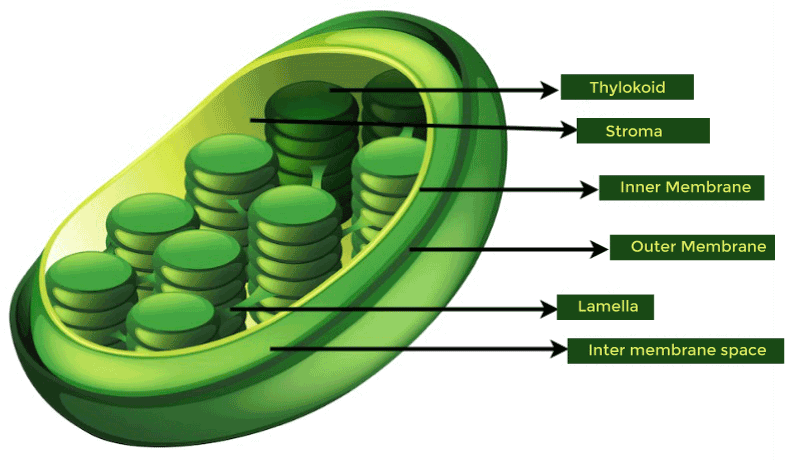
3. EtioplastsEtioplasts are certain intermediate plastid types that are mostly present in seedlings that have been developed in darkness. They are readily discovered in seedlings that emerge from the earth in their natural environment. Because they halt the development of photosynthetic chemicals and structures that are unnecessary in the dark, they are also regarded as the temporary stage of development for chloroplasts. In etioplasts, a single well-arranged paracrystalline prolamellar body and tubular prothylakoids are typically formed. These are interspersed with numerous small plastoglobules that contain high concentrations of carotenoids, especially lutein and violaxanthin, which aid in accelerating the transition to chloroplasts. Leucoplasts and Their Derivatives The white structures that makeup leucoplast are distinctive. They are often present in tissues with storage needs that are not photosynthesis-related. Leucoplast can now be classified in more depth because of improvements in microscopy technology and a rise in detection methods. Three kinds of white plastids?amyloplasts, proteinoplasts, and elaioplasts?are further classified as subtypes of leucoplasts, with the exception of undeveloped proplastids. Amyloplasts Amyloplasts are distinguished by high-density starch-storing starch granules. Free fatty acids, lysophospholipids, lysophosphatidylcholine, and lysophosphatidylethanolamine are only a few of the lipids that are present in the starch granules during the creation of amyloplast membranes. Amyloplasts are often found at low frequencies in a variety of tissues, including leaves, stems, and roots for temporal storage. They are also frequently found in sink tissues, such as seeds, fruits, tubers, and roots for carbon storage. It's intriguing that the amyloplasts are not always stable; for instance, in the leaves of Arabidopsis, starch accumulation and loss are very dynamic, occurring in a daily cycle as a result of photosynthetic activity or its absence. Amyloplasts often coexist in cells alongside other forms of plastid, unlike other types of plastid. Amylochromoplasts, a form of combination plastid that stores starch granules and carotenoid crystals in the same plastid, were discovered in the tissues of species such as the winter squash, peach palm fruit, and sweet potato tuber. Granules of starch may also be found within various plastid types, such as chloroplasts. The amyloplast from Arabidopsis roots was said to provide storage purposes as well as contribute to gravitropism signalling. Proteinoplasts Specific instances of protein bodies occurring in plastid structures?typically in the cytosolic region?are known as proteinoplasts. At various stages of plastid development, proteinoplasts can typically be found in a wide variety of cell types. Proteinoplasts are believed to have a function in protein storage due to their location and composition. Furthermore, the tobacco root's proteinoplasts displayed significant oxidase activity, suggesting that they may serve a particular purpose. Chromoplasts Because they store a lot of carotenoids, chromoplasts exhibit a variety of colours, each of which is caused by a different kind of carotenoid. The concentrated carotenoids created and stored in plastoglobules, which are hydrophobic structures, are concentrated during chromoplast growth and eventually form globular, spherical, coiled-shaped carotenoid crystals. The lipid production, storage, and cleavage processes are all carried out by the plastoglobules, which are lipoprotein particles linked to thylakoids by a half-lipid bilayer. In reproductive tissues, these colourful plastids with highly developed plastoglobules are utilised to entice pollinators and seed dispersers or to store carotenoids and hydrophobic compounds. 4. GerontoplastsGerontoplasts are plastids modified for recycling that are formed from chloroplasts and are often observed during senescence processes or under stressful conditions. Chloroplast disintegration and the recycling of their nutrients are crucial for plant life since they contain up to 80% of the leaf's nitrogen pool. Furthermore, autophagy, Senescence Associated Vacuoles (SAV), and Chloroplast Vesiculation (CV) have all been reported to be involved in the degradation of chloroplast proteins. Plastids experience a series of alterations in their ultrastructure once the senescence process begins. The properties of gerontoplasts at the start of senescence are difficult to pin down. However, a few distinctive traits have been noted. First off, gerontoplasts lack starch granules, perhaps because they are unable to sustain the photosynthesis that replenishes the starch on a regular basis. The degradation of their chlorophyll and thylakoid structures is the second factor. Third, their plastoglobules' size and quantity are growing, most likely as a result of the buildup of lipophilic materials from broken-down lipid structures and hydrophobic contents. InheritanceThe plastids often come from only one parent in plants. The majority of gymnosperms acquire plastids from the male pollen, while angiosperms typically get plastids from the female gamete. Additionally, algae only receive plastids from one parent. Thus, the other parent's plastid DNA is completely lost. Plastid DNA is inherited in normal intraspecific crossings (producing normal hybrids of one species) as a tight, one-parent inheritance. The inheritance of plastids, however, is more irregular in interspecific hybridizations. There are several cases of flowering plant hybrids that include father-specific plastids, despite the fact that plastids in interspecific hybridizations are typically inherited maternally. Alfalfa (Medicago sativa), among the 20% of angiosperms, often exhibits biparental plastid inheritance. Repair and Damage to DNAGrowing maize seedlings are more vulnerable to damage to their plastid DNA. In oxidative conditions produced by photo-oxidative reactions and photosynthetic/respiratory electron transfer, the DNA is harmed. While some DNA molecules undergo repair, unrepaired damage to DNA causes it to degrade into non-functional fragments. DNA repair proteins are encoded by the nuclear genome of the cell. Still, they may be transported to plastids, where they repair the DNA of the plastid to ensure genome stability/integrity. For instance, to preserve the integrity of the plastid genome, the DNA mismatch repair protein Msh1 interacts with the recombinational repair proteins RecA and RecG in the chloroplasts of the moss Physcomitrella patens. Summary and PerspectiveThe analysis of several representative samples demonstrated that sophisticated regulation of plastid interconversion is a crucial component for many agronomic characteristics. Not only has the development and interconversion of plastid affected the production potential of various plant-derived products, such as starch, fat, protein, and secondary metabolites. Despite the fact that environmental conditions are limited and plant demands are constantly rising, studying plastids may be the key to finding a breakthrough solution. It also provides scientific evidence in support of plastid research, including core regulators for candidates for molecular modification. Last but not least, candidate genes from the concisely described molecular pathway for plastid interconversion may be used as target genes to enhance a variety of agronomic qualities that are connected to plastid interconversion.
Next TopicPolymerization Definition
|
 For Videos Join Our Youtube Channel: Join Now
For Videos Join Our Youtube Channel: Join Now
Feedback
- Send your Feedback to [email protected]
Help Others, Please Share








